Gary Bauchan1 and Stephanie Greene2
1USDA-ARS
Soybean & Alfalfa Research Lab.
Beltsville, MD 20705
Phone: (301) 504-6649
E-mail: BauchanG@ba.ars.usda.gov
2USDA-ARS
Washington State University Irrigated Ag.
Research and Extension Center,
Prosser, WA 99350
Phone: (509) 786-9265,
E-mail: sgreene@ars-grin.gov.
Chapter 1 - The Genus Medicago
Introduction
Alfalfa ...........................................................................................................................
2
Production ................................................................................................................
3
Highlights of Alfalfa Production .................................................................................
5
Value ........................................................................................................................
5
Annual Medics ................................................................................................................
6
What are medics? .......................................................................................................
6
Uses of medics ............................................................................................................
7
Chapter 2 - Genetic Vulnerability in Alfalfa .............................................................................
7
Introduction ...................................................................................................................
7
Origins of Alfalfa Cultivars (1830 - 1977) .......................................................................
7
List of Diseases, Insects and Nematodes of Alfalfa ......................................................
8
Alfalfa Genetic Vulnerability ...........................................................................................
8
Current Status (1985 - 2000)..........................................................................................
8
List of Threatened Medicago species ................................................................................
10
Chapter 3 - Alfalfa Breeding .....................................................................................................
10
Introduction .....................................................................................................................
10
Alfalfa Crop Germplasm Committee (ACGC)...................................................................
12
Formation of ACGC .....................................................................................................
12
Accomplishments of ACGC ..........................................................................................
13
Locations Conducting Germplasm Research .....................................................................
14
Chapter 4. National Plant Germplasm System Medicago Germplasm Collection ....................
16
Introduction ....................................................................................................................
16
History .............................................................................................................................
16
Core Subsets ....................................................................................................................
18
Introduction .....................................................................................................................
18
Perennial Medicago Core Subset ..................................................................................
18
Annual Medic Core Subset ..........................................................................................
19
Rhizobium Collection ..................................................................................................
20
Assessing Collection for Current Gaps .............................................................................
21
Primary gene pool ...........................................................................................................
21
Obsolete and Current Cultivars ....................................................................................
22
United States ...............................................................................................................
23
Other Nations .............................................................................................................
23
Landraces & Primitive Cultivars ..................................................................................
23
Wild Medicago subsp. sativa .......................................................................................
25
Medicago sativa Complex ...........................................................................................
25
Secondary Gene Pool .....................................................................................................
26
Tertiary gene Pool ..........................................................................................................
27
Acquisition Priorities ......................................................................................................
27
Cultivated Medics .....................................................................................................
27
Non-cultivated Medicago species ...............................................................................
27
Collection Evaluations ....................................................................................................
28
Perennials .................................................................................................................
28
Annual Medics ...........................................................................................................
29
Germplasm Enhancement ................................................................................................
29
NPGS Germplasm Collection Use 1993-1997 ................................................................
30
Accession Availability ................................................................................................
31
Collection Preservation ...............................................................................................
31
Appendix 1. Taxonomic listing of Medicago species, geographic distribution,
number of accessions, and conservation status ...................................................................
33
Appendix 2. Evaluation status of the perennial species ..........................................................
37
Appendix 3. Evaluation status of the annual species ..............................................................
42
Appendix 4. Germplasm enhancement proposal for development of national and regional
germplasm pools ................................................................................................
46
References ...............................................................................................................................
50
Compilation of Recommendations ..........................................................................................
51
Table 1. List of cultivated Medicago species their common
name, usage and growth cycle........ 1
Table 2. Alfalfa and all-hay production by states in 1998
.......................................................... 3
Table 3. List of institutions and locations were germplasm
research is taking place.................. 14
Table 4. Alfalfa germplasm collection trips made by U.S.
scientists in the last 100 years ........... 16
Table 5. Number of cultivars and country of origin represented
in the NPGS collection............. 22
Table 6. Number of accessions representing wild relatives
(primary gene pool) of alfalfa
in the NPGS collection................................................................................................
26
Figure 1. All hay production (Acres & Production) 1865-1998................................................
4
Figure 2. Alfalfa hay production (Acres & Production) 1940 - 1998 ........................................
4
Figure 3. Germplasm sources us in alfalfa cultivars (All dormancy classes) 1986
- 1999 .......... 9
Figure 4. Germplasm sources us in alfalfa cultivars (Dormant) 1986 - 1999 .............................
9
Figure 5. Germplasm sources us in alfalfa cultivars (Non-dormant) 1986 - 1999
..................... 10
Figure 6. Distribution of landrace germplasm held in the NPGS collection ................................
24
Figure 7. World distribution of cultivated germplasm held in the US collection.......................
24
Figure 8. World distribution of wild Medicago sativa subsp. sativa germplasm
held in
the NPGS collection...................................................................................................
25
Figure 9. Number of seed requests (1993 to 1995) ..................................................................
31
Alfalfa acreage is at 23.6 million acres and is predicted to maintain or drop slightly in acreage primarily due to the increase productivity of each acre.
Alfalfa is the best animal feed for ruminant livestock and horses and it ranks 4th among all U.S. agricultural crops grown with an annual production value of $8.1 billion . When the value of animal products is added to the value of alfalfa hay, the dollar value of alfalfa reaches $145 billion annually. Thus far exceeds the combined value of all other high value crops.
Annual medics have potential uses in sustainable agriculture systems, however, additional research is needed to discover their niche in the U.S.
The general impression has been that crop genetic vulnerability was not a large concern due to the autotetraploid nature of Medicago sativa subsp. sativa, the high level of outcrossing, and the diversity of germplasm sources utilized in the development of alfalfa cultivars. However, a survey of the cultivars released over the past 14 years has shown a narrowing of the germplasm utilized.
Development of alfalfa cultivars has shifted from the public sector to private companies over the past 60 years. Today, virtually all commercial alfalfa cultivars are developed by private companies.
The USDA-ARS breeding effort has shifted towards germplasm collection, evaluation, enhancement and development as well as fundamental classical and molecular studies on alfalfa and related species.
The University's have also shifted towards germplasm evaluation, enhancement and development, however, a few public breeders continue to develop special use cultivars of alfalfa. The University's have also been involved in fundamental studies of alfalfa particularly in the area of molecular biology.
Organizations involved in germplasm development include federal, state, and private companies. The North American Alfalfa Improvement Conference continues to be the conduit through which scientific information is exchanged between public organizations and private companies for the benefit of the alfalfa crop.
1. Collection maintenance
2. Germplasm collection
3. Germplasm evaluation
4. Germplasm enhancement
Although the seed regeneration of almost all the collection has been completed, the need remains to a) regenerate accessions requiring special care/handling, b) obtain baseline seed viability data, c) store original seed and regeneration seed lots at -18 C, d) deal with backlog of "W6" and NSL accessions (ie. should they go into collection?)and duplicate accessions, and e) verify our regeneration procedures are adequate. Without the continued vigilance in maintaining our current collection, future collections, evaluations and enhancements would be of little value.
The greatest needs for the collection are for the wild perennial species in the secondary and tertiary germplasm pools, especially those species which are considered threatened.
Countries where future collections should occur include Georgia, Crimea and Southern Caucasus mountain regions of the former Soviet Union, the Himalayan region of India, south and north east regions of China, Sudan, Egypt and the Arabian Peninsula, South Africa, Northern Kazakhstan, and adjacent Central Asian countries (i.e. Kyrgistan, Uzbekistan, Afghanistan), Iran and Iraq.
A set of descriptors for all accessions, especially all alfalfa accessions needs to be completed. The currently established core subsets should be reevaluated once descriptor data has been obtained on the entire collection. Accession classification based on molecular markers needs to be accomplished and molecular data compared to classifications based on morphological and ecogeographic descriptors. In addition, evaluations which can broaden the use of alfalfa need to be carried out.
National and regional germplasm pools should be developed.
A Compilation of the Specific Recommendations is Located on Page 51.
The genus Medicago is a member of the Legume family. According to Small and Jompe (1989) the genus contains 30 perennial species and 60 annual species (Appendix 1). About 15 species are cultivated for various uses (Table 1.):
Alfalfa (Medicago sativa L. sensu lato) is a perennial species and the most important forage legume in North America. The annual species, referred to as medics, serve as important components in pastures in regions with Mediterranean climates.
Table 1. List of cultivated Medicago species their common name, usage and growth cycle.
| Cultivated species of Medicago |
Common name |
Use | Growth Cycle |
|---|---|---|---|
| Medicago arabica (L.) Huds. | Spotted bur clover, spotted medic | Animal forage | Annual |
| Medicago arborea L. | Tree alfalfa, tree medic | Animal forage, ornamental | Perennial |
| Medicago italica (Mill.) Fiori |
Disc Medic | Animal forage, soil improvement, companion crop | Annual |
| Medicago littoralis Rohde ex Loisel. | Strand Medic | Animal forage, soil improvement, companion crop | Annual |
| Medicago lesinsii | Animal forage, soil improvement, companion crop | Animal | |
| Medicago lupulina L. | Black medic | Animal forage, soil improvement, companion crop | Annual |
| Medicago minima (L.) Bartal. | Little bur clover | Animal forage |
Annual |
| Medicago murex Willd. Annual | Animal forage, soil improvement, companion crop | Annual | |
| Medicago orbicularis (L.) Bartal. | Button clover | Animal forage, soil improvement, companion crop | Annual |
| Medicago polymorpha L. | Toothed bur-clover, Toothed medic, California bur-clover | Animal forage, soil improvement, companion crop | Annual, biennial, perennial |
| Medicago rigidula (L.) All. | Tifton bur-clover, Tifton medic | Animal forage, soil improvement, companion crop | Annual |
| Medicago rugosa Desr. | Gama medic | Animal forage, soil improvement, companion crop | Annual |
| Medicago sativa nothosubsp.
varia (Martyn) Arcang. |
Sand lucerne , variegated lucerne |
Animal forage | Perennial |
| Medicago sativa subsp. falcata (L.) Arcang. | yellow-flower alfalfa, sickle alfalfa, yellow Lucerne | Animal forage | Perennial |
| Medicago sativa subsp. sativa | Alfalfa , Lucerne | Animal fodder and forage, soil improvement, human food-sprouts, medicinal-tea | Perennial |
| Medicago scutellata (L.) Mill. | Snail medic | Animal forage, soil improvement, companion crop | Annual |
| Medicago truncatula Gaertn. |
barrel medic | Animal forage, soil improvement, companion crop | Annual
|
Alfalfa, called the "Queen of the Forages," is the fourth most widely grown crop in the United States. Alfalfa hay is used primarily as animal feed for dairy cows but also for horses, beef cattle, sheep, chickens, turkeys and other farm animals.
Alfalfa is recognized as one of the most important crops to U.S. agriculture due to its ability to:
In addition to the traditional uses of alfalfa as an animal feed, alfalfa is
beginning to be used as a bio-fuel for the production of electricity, bioremediation
of soils with high levels of nitrogen, and as a plant factory for the production
of industrial enzymes such as peroxidase, alpha-amylase, cellulase, and phytase.
Alfalfa is also consumed directly by humans as alfalfa sprouts and alfalfa juice
is found in some health food products. Plant molecular biologist use alfalfa
as a model crop due to it ability to grow entire plants from single cells and
it's ability to be genetically transformed thus allowing improvements of the
crop through gene insertion.
Alfalfa is grown in all 50 states over a wide range of climatic conditions
from fall dormant types in the Midwest and Northeast where winter survival is
key, and non-dormant types in the Southwest U.S. where extensive growth is needed
from October to March. There are 23.6 million acres of alfalfa cut for hay with
an average yield of 3.5 tons per acre. Alfalfa is sometimes grown in mixtures
with forage grasses and other legumes. Since the introduction of alfalfa into
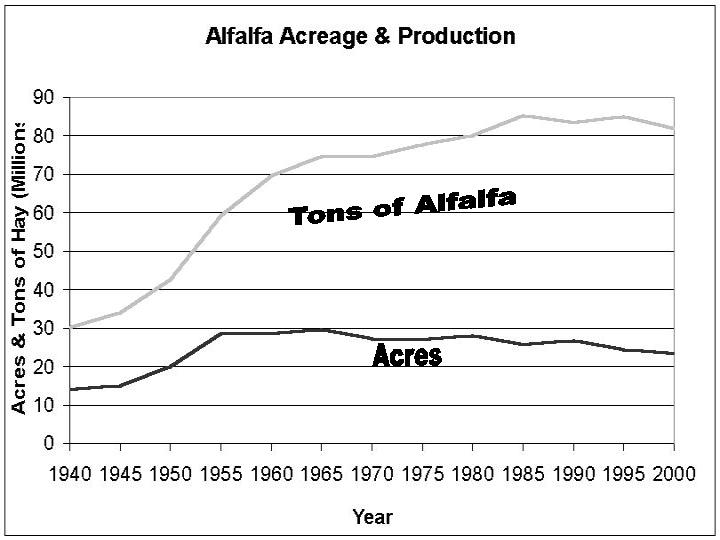
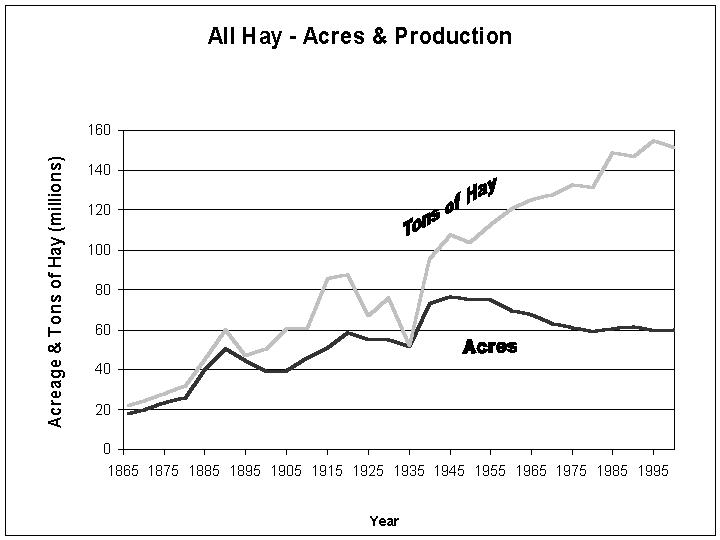
the U.S. the productivity of forages has dramatically increased (Figure 1).
The acreage of all hay harvested including alfalfa is 60.8 million per year.
The production of alfalfa hay on a large scale began around 1919 in the U.S.
and has grown from 14 million acres in 1940 to it's highest number of acres
of nearly 30 million acres in 1965. Since 1965 the total number of acres of
alfalfa has decreased slightly, however, the productivity of alfalfa fields
has been steadily increasing (Figure 2). Over the past 10 years the total number
of acres has decreased from 26.7 million acres in 1987 to 23.6 million acres
in 1997, however, the production has increased from 2.59 tons per acre with
a total of 69.3 million tons to 3.47 tons per acre with a total of 82 million
tons. The increased productivity of alfalfa over the years can be attributed
to better management practices, genetic improvement of cultivars, and the increased
use of mechanization for harvesting alfalfa.
Table 2. Alfalfa and All-Hay production by states in 19981.
| Region |
States |
Alfalfa Production Tons X 106 |
Alfalfa Acreage Acres X 106 |
All Hay Production Tons X 106 |
All Hay Acreage Acres X 106 |
|---|---|---|---|---|---|
| East |
CT, DE, MA, ME, MD, NC, NH, NJ, NY, PA, RI, VA, VT |
4.5 |
1.7 |
12.6 |
6.1 |
| South East |
AL, FL, GA, LA, MS, SC |
0.0 |
0.0 |
7.0 |
2.9 |
| Central |
AR, IL, IN, KY, MI, MO, OH, TN, WV |
11.8 |
3.4 |
33.2 |
13.5 |
| North Central |
IA, MN, MT, NE, ND, SD, WI |
29.9 |
11.6 |
41.3 |
19.6 |
| Great Plains |
CO, KS, OK, TX |
9.4 |
2.2 |
27.0 |
11.0 |
| Intermountain |
ID, UT, WY |
8.6 |
2.2 |
10.4 |
3.3 |
| South West |
AZ, NM, NV |
4.1 |
0.7 |
4.8 |
1.1 |
| North West |
CA, OR, WA |
11.5 |
1.9 |
15.3 |
3.3 |
| Totals |
|
79.8 |
23.7 |
151.6 |
60.8 |
1 Statistics obtained from the 1998 USDA - National Agricultural Statistics Service.
1850 Introduction of non-dormant alfalfa's from South America into southwesternUS
(Chilean)
1901 First field trials of dormant alfalfa in Minnesota (Grimm)
1940 Shortage of winter-hardy seed affected acreage planted.
1942 First two bacterial wilt-resistant cultivars are released. (Ranger and
Buffalo)
1943 Introduction of mechanized hay conditioners.
1948 Forced-air hay dryers introduced and wilted silage generally recommended.
1953 First high winter-hardy bacterial wilt resistant variety released. (Vernal)
1954 First variety of nematode and bacterial wilt resistant variety is released
(Lahontan)
1955 Direct-cut silage increases in popularity associated with development of
new equipment.
1957 Release of the first spotted aphid resistant varieties. ( Moapa and Zia).
1958 Alfalfa seed industry initiates research on alfalfa breeding.
1963 Release of Flemish type alfalfa with bacterial wilt resistance. (Saranac)
1968 Release of a moderately resistant variety to the alfalfa weevil. (Team)
1968 First variety released with pea aphid resistance. (Kanza)
1970 Beginning of use of hay and silage preservatives.
1970 Release of a variety with resistance to four biotypes of the spotted alfalfa
aphid. (Hayden)
1972 First variety released with Phytophthora root rot resistance. (Agate)
1973 First variety released with Anthracnose resistance (Arc)
1976 First variety released with blue aphid resistance (CUF-101)
1979 Cultivated alfalfa at the diploid level developed for basic genome studies
(CADL)
1980 First release of multifoliolate leaf variety. (Multileaf)
1981 Release of five varieties with resistance to Verticillium wilt resistance.
(Apollo II, DK-135, Trumpetor, WL 316 and Vernema)
1982 Ten-ton hay yield barrier broken without irrigation in East Lansing, Michigan
1983 USA hay quality standards developed for ADF, DDM, CP and DM..
1986 Release of a variety with increased N2 capabilities for short-term crop
rotation. (Nitro)
1989 First germplasm release of highly regenerable tissue culture type. (Regen
S)
1991 Release of a grazing tolerant alfalfa. (Alfagraze)
1991 Release of the first high quality alfalfa ( WL322HQ)
1997 First variety released with potato leafhopper resistance (Trailblazer)
Alfalfa is primarily grown on the farm in which it is consumed, thus it is
difficult to estimate the actual value of alfalfa. Alfalfa hay is used primarily
as animal feed with an estimated value of $ 8.1 billion. Alfalfa is also grown
in mixtures with forage grasses and other legumes. The estimated value of all
hay is $13.4 billion. The average cost of alfalfa hay is $102.50 per ton. Alfalfa
meal and cubes are exported to other countries with a value of $49.4 million
to the U.S. economy When the value of animal products estimated at $132 billion
from products such as milk, cheese, butter, meat, wool, etc. is added to the
value of hay, the value of alfalfa reaches the $145 billion level. This far
exceeds the combined value of all other high value crops.
Alfalfa seed is primarily grown in the western areas of the U.S. primarily in
the states of California, Idaho, Nevada, Oregon, Wyoming, and Washington. The
total U.S. production of alfalfa seed in 1999 was 115 million pounds, with an
average price of $190 per 100 pounds of seed. Thus, the estimated value of alfalfa
seed in the U.S. is $218.5 million. A fringe benefit to the production of alfalfa
seed is the production of honey from bees. In the U.S., $147.7 million dollars
worth of honey is produced each year.
Alfalfa is also directly consumed by humans in the form of alfalfa sprouts. According to the International Sprout Growers there are approximately $250 million dollars worth of sprouts sold annually in North America. Alfalfa juice is used in some health food products with an undefined economic value.
Annual Medicago species, referred to as medics, are excellent candidates for use in sustainable agriculture systems such as pastures and cover crops. Medics are native to semiarid areas around the Mediterranean Sea and have long been used in unimproved pastures in the region. Medics are now widely distributed throughout the world, largely in areas with mild, rainy winters and alkaline soils. Medics are among the principal legumes used in the Australian ley farming system, and constitute over 50 million hectares of the Australian agriculture zone (Crawford et al., 1989). Medics have been most successful in cereal-legume crop rotation systems in southern Australia where winter rainfall averages between 250 and 500 mm. The species most widely grown in Australia are: M. littoralis Rohde ex Lois., M. polymorpha L., M. rugosa Desr., M. scutellata (L.) Miller, M. italica (Miller) Fiori, and M. truncatula Gaerth.
The agronomic potential of the medics has been investigated and several different systems have been developed. 1) The medics can be used as permanent pasture; 2) medics can be used as a green manure crop due to it's ability to fix nitrogen and as a cover crop to prevent soil erosion; 3) annual medics can be used as a companion crop to control weeds, add organic matter, lower soil temperatures and provide fall residue after harvest. Various crops have been utilized including corn, soybeans, small grains, sunflower, squash and some herbs; 4) vineyards and fruit orchards have used annual medics in the alleyways to provide ground cover for soil erosion, an environment for beneficial insects, and a stable track for equipment; and 5) use of annual medics as wildlife habitat for deer, quail and water fowl have been initiated. Medics do have potential uses in sustainable agriculture systems, however, additional research is needed to discover their niche (Bauchan, 1999).
The general impression has been that crop genetic vulnerability was not a large concern due to the autotetraploid nature of Medicago sativa subsp. sativa, the high level outcrossing, and the diversity of germplasm sources utilized in the development of alfalfa cultivars between 1930 and 1977 (Barnes 1977). Two decades have past since this initial assessment was made, it seems prudent to reassess the present day status of the genetic vulnerability in alfalfa.
Alfalfa originated in Vavilov's "Near Eastern Center"--Asia Minor, Transcaucasia, Iran and Turkistan. In the wild, M. sativa and related perennial species are found throughout Eurasia and as far north as Siberia. Alfalfa spread from its center of origin into Europe, North Africa, Arabia and eventually South America with invading armies, explorers, and missionaries as feed for horses and other livestock. In 1736, European colonists brought alfalfa to the eastern U.S. where the crop was referred to by its European name, lucerne. These introductions generally were not successful, except for a few planted on well-drained limestone soils. Alfalfa was well suited to the dry climates and irrigated soils of the western U.S., where it was introduced from Mexico by Spanish missionaries as early as the 1830s. Alfalfa eventually spread eastward to the Intermountain region and the southern Great Plains. Movement into areas with severe winters was limited by the lack of winter hardiness in the primarily Spanish-derived (non-dormant) germplasm. The introduction of four winter-hardy (dormant) types (cv. Grimm, cv. Ontario Variegated, cv. Baltic, and cv. Cossack) from northern Europe to the North Central States between 1858 to 1910 allowed successful alfalfa culture in the colder and more humid areas of the Midwestern and Northeastern U.S.
Before 1925, most alfalfa breeding efforts in North America were directed toward selecting strains that were more winterhardy. During the next 30 years, emphasis was placed on developing cultivars that combined winterhardiness and resistance to bacterial wilt. During the late 1950s, the emphasis was placed on developing cultivars resistant to other diseases and several insect pests (Barnes et al., 1988). Beginning in the 1950's there was an emphasis on breeding alfalfa with multiple pest resistance through recurrent selection breeding schemes. Most modern varieties of commercial alfalfa have moderate to high levels of resistance to the major disease, insect and nematode pests. Scientists belonging to the North American Alfalfa Improvement Conference have developed standardized test procedures for screening alfalfa for the following diseases, insects and nematodes.
Anthracnose
In assessing the genetic vulnerability of alfalfa in 1977, Barnes credited nine distinct germplasm sources which were introduced into the U.S. from different regions of the world, in the development of modern alfalfa varieties. They are described in descending order from the most winter hardy (most fall dormant) to the least winter hardy (least fall dormant): ‘Falcata', ‘Varia', ‘Turkistan', ‘Flemish', ‘Chilean', ‘Peruvian', ‘Indian", and ‘African' (Barnes et al. 1977). The 1977 report concluded that a trend toward increased genetic diversity in alfalfa cultivars, spurred largely by breeding efforts begun in the 1950s, and the inherent genetic heterogeneity of the species had made alfalfa less vulnerable to catastrophic loss than it was prior to the 1950's. However, the report also stated that "the broad use of the recognized germplasm in most varieties is cause for concern about future yield improvements" (Barnes 1977).
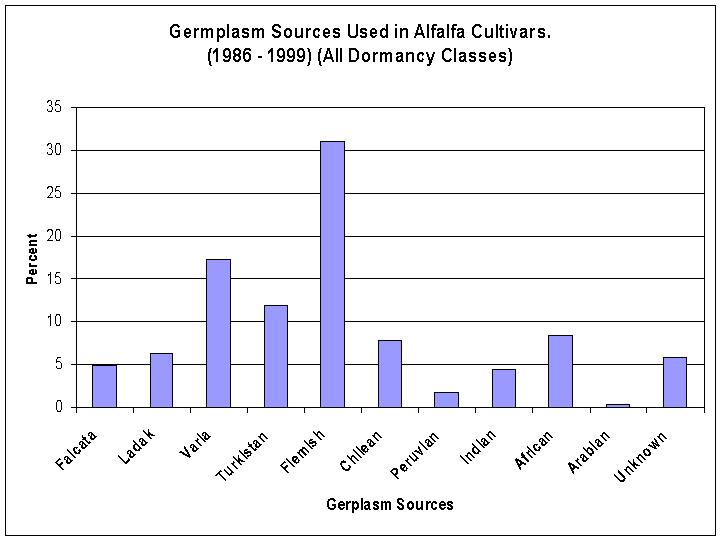
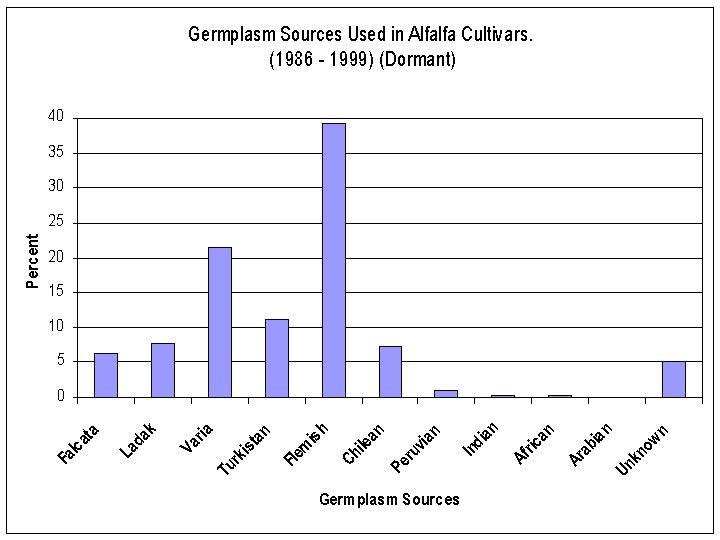
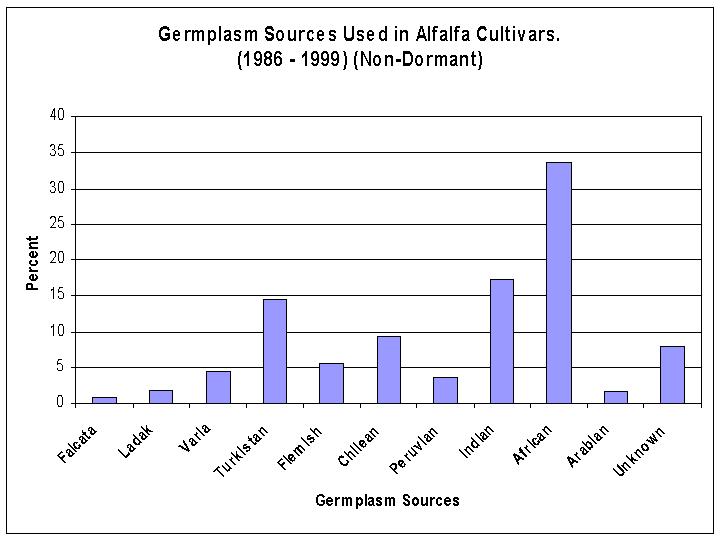
Over the past 15 years (1985-2000) a majority (76%) of the cultivars released have been fall dormant type alfalfas. There are three germplasm sources that have been used extensively, Flemish (31%), Varia (17%) and Turkistan (11%) (Figure 3). Within the dormant alfalfas, most of the selections have been made from two germplasm sources, Flemish (39%) and Varia (21%) (Figure 4). Whereas, within the non-dormant types of alfalfa most of the selections have been made from three germplasm sources, African (34%), Indian (17%), and Turkistan (14%) (Figure 5). Research begun in the 1980's increased the use of plant introductions which were wild perennial relatives of alfalfa. Three perennial species, M. glandulosa, M. glutinosa, and M. prostrata are recognized as the sources of glandular hairs which have been transferred to alfalfa to provide tolerance to the potato leafhopper.
There is a need to broaden the germplasm base used in alfalfa cultivars released by the alfalfa seed industry. The introduction of wild perennial germplasm and very non-dormant types from the Saudi Arabian Peninsula are examples of the beneficial attributes available.
Enhanced support for germplasm evaluation and enhancement projects would encourage the use of a broader germplasm pool by plant breeders which would lead to more diverse alfalfa cultivars.
The following Medicago species have been listed by the World Conservation Monitoring Center as having a world status of vulnerable or rare:
M. cancellata*
M. citrina
M. cyrenaea
M. pironae*
M. rhodopea*
M. shepardii
M. strasseri
* Note that this list includes three species considered to be wild relatives of cultivated alfalfa.
The first breeding efforts in the U.S. date back to the 1850's for non-dormant
alfalfas and the early 1900's for dormant types of alfalfa. Most alfalfas grown
in the early years were selections of the best materials imported from Europe
and Russia. In the Midwest in the early 1900's the emphasis was the selection
for winter hardiness. From alfalfa's initial introduction until 1955, about
33 recognized alfalfa cultivars or regional strains were grown in the U.S. and
Canada. About half of these were directly introduced from Europe, Asia or the
Near East. Between 1958 and 2000, organized alfalfa breeding expanded tremendously
and the number of recognized cultivars increased to about 400. From the 1900's
until the 1950's almost all of the new alfalfa cultivars were developed by the
USDA and public university programs. The goal of these programs was primarily
to increase winter hardiness and the development of bacterial wilt resistance.
Starting in the 1950's the alfalfa seed industry began. Since their inception,
private alfalfa breeding programs in the U.S. have been responsible for the
rapid increase in numbers of recognized alfalfa cultivars. During the period
between 1955 and 1960, approximately 20% of cultivars were released from private
breeding programs. This proportion increased to more than 95% between 1985 and
1990 and nearly 100% between 1990 and 2000. Private breeding programs increased
in number, size and sophistication from about 1965 to 1985. During the late
1980s, competition for market share caused several mergers within the private
sector. At present, there are about 8 commercial alfalfa breeding programs with
annual research expenditures ranging from nearly $100,000 with 1.0 scientist-year
(SY) to more than $2 million with five or more SYs. The rate of release of new
alfalfa cultivars in the U.S. has increased steadily over the last 90 years.
About one cultivar was released every three years between 1901 and 1940; one
each year between 1941 and 1960; about 17 per year between 1981 and 1985; about
30 each year between 1985 and 1990; about 60 per year between 1990 and 1995,
and about 100 per year between 1996 and 2000.
Increased breeding by industry was accompanied by a reduction in the number
of public breeding programs, and by a change in the direction of public research
from breeding new cultivars to developing new breeding procedures and improved
germplasm (Barnes et al., 1988). The transition from applied alfalfa breeding
to more fundamental research within public agencies contributed to the development
of several comprehensive research efforts. Examples of research areas include:
More generally, public researchers working with alfalfa also continue to provide valuable knowledge in plant genetics, physiology, and growth, including resistance to biotic and abiotic stresses. Public research in cooperation with industry partners has lead the way in developing potentially new uses of alfalfa for a bio-fuel for the production of electricity, bioremediation of soils with high levels of nitrogen, and as a factory for the production in bioreactors of industrial enzymes.
In discussing research directions for the 1990s and beyond, the Alfalfa Crop Germplasm Committee (ACGC) listed the following as new uses or value-added traits that could be developed in alfalfa (Minutes of the Alfalfa CGC, October 9-10, 1990, Kansas City, MO):
The increased commitment to alfalfa improvement in the private sector has been associated with a concomitant decline in the number of public alfalfa researchers. The wide acceptance of the best proprietary cultivars by the 1980s confirms the efficacy of private breeding programs, and suggests that an appropriate balance has been reached between fundamental public research and more applied private research. However, the balance between public and private programs may now be in danger because of reductions in support for public alfalfa research programs. The termination of a number of public alfalfa breeding and genetics programs also means that many breeder's germplasm collections may be subject to loss.
The Alfalfa Crop Germplasm Committee consists of 21 members representing federal, state and private interests. There are representatives of several scientific disciplines including: breeding, genetics, cytogenetics, entomology, pathology, physiology, microbiology, and molecular biology. In addition there is geographic representation of breeders from the major alfalfa growing areas of the U.S.
The Alfalfa Crop Germplasm Committee address critical issues facing the National Plant Germplasm System (NPGS) especially related to the genus Medicago. These include:
The Alfalfa Improvement Conference (now the North American Alfalfa Improvement Conference - NAAIC) has since it's inception in 1934 been concerned with alfalfa germplasm. In the early days of the conference there was need for germplasm exchange among scientists, seed production of experimental strains in the western U.S., development of a standardized system of note taking, and testing alfalfa cultivars for disease and insect pests, yield, and persistence. Several committees grew out of the Alfalfa Improvement Conference which were related to germplasm, they included: variety testing, release of new cultivars, available breeding lines, lists of cultivar descriptors, development of standardized tests for characterizing pest of alfalfa, and the National Alfalfa Variety Review Board. The Alfalfa Crop Germplasm Committee was formerly organized in 1969 as the Germplasm Committee of the Alfalfa Improvement Conference.
1970 Developed a program for carefully controlled seed increase of the Medicago
collection using isolation cages. Previous to this the germplasm collection
was increased using open pollination.
1977 Developed the statement on the genetic vulnerability of alfalfa.
1977 Developed a plan to produce seven (7) regional pools of alfalfa germplasm
as a means of preserving germplasm and evaluating germplasm for suitability
of growth in a region.
1978 Alfalfa is included as one of the first crops to be included in the Germplasm
Resources Information Project (now the Germplasm Resources Information Network
- GRIN).
1979 Alfalfa seed increase was initiated in Reno, Nevada of at least one pound
per accession under isolation cages.
1980 Plant explorations were undertaken in Chile, Bolivia, Peru, Ecuador, Turkey
and old stands of alfalfa in the U.S. and Canada.
1981 Evaluation of the germplasm collection was initiated using standardized
tests for 9 diseases, 5 insects, and 3 nematodes. Data was entered into GRIN.
1982 Germplasm collection trips were successful in collecting 561 accessions
in the USSR and Morocco.
1984 The Karl Lesins Collection from Canada composed of over 3000 annual and
600 perennial accessions were added to the U.S. germplasm collection.
1985 Germplasm enhancement program developed.
1986 Seed increase project is transferred to Prosser, Washington.
1987 Committee membership expanded beyond only plant breeders to include representation
of various disciplines including, cytogenetics, entomology, pathology, microbiology,
and molecular biology.
1987 Transfer of the Medicago collection from Ames, IA to Pullman, WA.
1988 Annual medic seed increase initiated in Riverside, CA.
1990 Evaluations of the germplasm collection using standardized tests for agronomic
traits such as fall dormancy, winter and frost injury, forage quality, salt
tolerance, and tolerance to acid soils. Data entered into GRIN.
1991 Core subsets were developed for perennial and annual Medicago species
as well as Rhizobium. Data are entered into GRIN.
1993 Medicago Germplasm Newsletter published.
1994 Majority of the germplasm collection has been evaluated for 35 agronomic
and pest resistance traits. Data are available in GRIN.
1995 Curator of the Medicago collection is employed.
1996 Plant explorations were undertaken in Mongolia and North Caucasus Mountains
of Russia.
1997 Seed increase of alfalfa collection under caged conditions largely complete.
Increase of annual medic collection largely completed.
1998 Passport and evaluation data in GRIN was updated and duplicate accessions
identified.
Table 3. List of Institutions and locations where germplasm research is taking place.
| Institution | Location | Agronomic | Physio-logical | Diseases | Insects | Nematodes | Bio-Tech | Pre-breeding | Cultivar Development |
|---|---|---|---|---|---|---|---|---|---|
| USDA-ARS |
Manhattan, KS |
X |
X |
X |
X |
X |
|||
| USDA-ARS |
Beltsville, MD |
X |
X |
X |
X |
X |
|||
| USDA-ARS |
St. Paul, MN |
X |
X |
X |
X |
X |
X |
||
| USDA-ARS |
Stoneville, MS |
X |
X |
X |
|||||
| USDA-ARS |
Logan, UT |
X |
X |
X |
X |
||||
| USDA-ARS |
Prosser, WA |
X |
X |
X |
X |
X |
|||
| University of Arizona |
Tucson, AZ |
X |
X |
X |
X |
||||
| University of California |
Davis, CA |
X |
X |
X |
X |
X |
X |
X |
|
| University of Georgia |
Athens, GA |
X |
X |
X |
X |
X |
|||
| Iowa State University |
Ames, IA |
X |
X |
X |
X |
||||
| Purdue University |
West Lafayette, IN |
X |
X |
X |
X |
||||
| Kansas State University |
Manhattan, KS |
X |
X |
X |
|||||
| University of Minnesota |
St. Paul, MN |
X |
|||||||
| New Mexico State University |
Las Cruces, NM |
X |
X |
X |
|||||
| Cornell University |
Ithaca, NY |
X |
X |
X |
|||||
| Ohio State University |
Columbus, OH |
X |
|||||||
| Oklahoma State University |
Stillwater, OK |
X |
X |
||||||
| University of Wisconsin |
Madison, WI |
X |
X |
X |
X |
||||
| University of Wyoming |
Laramie, WY |
X |
X |
X |
|||||
| ABI Alfalfa |
Ames, IA |
X |
X |
X |
X |
X |
X |
||
| Cal/West |
Woodland, CA |
X |
X |
X |
X |
X |
X |
||
| Dairyland Seeds |
Clinton, WI |
X |
X |
X |
X |
X |
X |
||
| Forage Genetics |
West Salem, WI |
X |
X |
X |
X |
X |
X |
X |
X |
| Great Plains |
Apex, NC |
X |
X |
X |
X |
X |
X |
||
| Pioneer Hi-Bred |
Johnston, IA |
X |
X |
X |
X |
X |
X |
X |
X |
| WL Research |
Evansville, WI |
X |
X |
X |
X |
X |
X |
X |
|
The National Plant Germplasm System Collection of Medicago germplasm is currently located at the Western Regional PI Station in Pullman, Washington. Stephanie Greene is the curator of the collection who can be contacted at the USDA-ARS, Washington State University Irrigated Agricultural Research and Extension Center, Prosser, WA 99350 Phone: (509) 786-9265, FAX: (509) 786-9370, e-mail: sgreene@ars-grin.gov. The current collection contains a total of 7,554 accessions with 4,108 perennial accessions representing 30 species and 3,446 accessions representing 60 annual species (Appendix 1).
The beginning of the U.S. Medicago germplasm collection traces back
to 1898. Accessions were collected directly in the field by plant explorers
and scientists or obtained from agricultural institutes, seed companies and
U.S. Embassy agricultural attaches. About 65 collection trips that focused on
alfalfa germplasm have been made in the last 100 years (Table 4). The majority
of accessions that are available today were received after the 1940s. During
the 1950s, 60s and 70s the collection grew by an average of 1400 accessions
per year. During the eighties, more than 4200 accessions were added to the collection.
During the nineties, the collection grew by 1400 accessions. The collection
currently contains over 7000 accessions representing 90 species from 94 countries.
Table 4. Alfalfa germplasm collection trips made by U.S.
scientists in the last 100 years
| YEAR |
COLLECTOR |
COUNTRY |
|---|---|---|
| 1898 | N.E. Hansen | Russia |
| 1899 | Lathrop, D. Fairchild |
Peru, Argentinia |
| 1899 | C. Sprenger | Italy |
| 1906 | N.E. Hansen |
Europe, Siberia, E. Asia |
| 1909 | N.E. Hansen | Central Asia |
| 1910 | F. Meyer |
Russia |
| 1924 | H.L. Westover | South America |
| 1925 | D. Fairchild | North Africa |
| 1925 | P.H. Dorsett |
China |
| 1926 | D. Fairchild | Northern Africa |
| 1929 | H.L. Westover | Russia |
| 1930 | H.L. Westover |
Turkestan, Europe |
| 1930 | P.H. Dorsett, W.J. Morse | China |
| 1936 | H.L. Westover; F.L. Wellman |
Turkey; Iran |
| 1937 | W. Koelz | Afghanistan |
| 1939 | W. Koelz | Afghanistan |
| 1940 | W. Koelz |
Iran |
| 1948 | J.R. Harlan | Turkey |
| 1951 | S.P. Kohli | India |
| 1951 | W.A. Archer |
Ethiopia |
| 1952 | R.K. Godfrey | Turkey |
| 1953 | H.S. Gentry | Afghanistan |
| 1954 | E.E. Smith | Afghanistan |
| 1954 | H.S. Gentry and E.E. Smith | Pakistan |
| 1955 | H.S. Gentry | Iran |
| 1955 | J.L. Stephens | E.Africa |
| 1956 | H.S. Gentry and H.A. Schoth | Europe |
| 1958 | H.S. Gentry and H.A. Schoth | Dinaric Alps |
| 1968 | W.H. Skarda | USSR |
| 1972 | D.R. Dewey | Iran |
| 1972 | J.L. Schwendiman | Turkey, Iran |
| 1973 | I. Forbes Jr., J.S. Gladstone | Morocco, Spain |
| 1975 | D.R. Dewey | Iran |
| 1975 | W.R. Lanford and B.L. Burson | Brazil |
| 1976 | P. K. Knowles | Afghanistan |
| 1977 | D. R. Dewey | USSR |
| 1977 | R.R. Smith; W.R. Landford | Greece, Italy; USSR |
| 1978 | D.R. Dewey and A.P. Plummer | USSR |
| 1978 | G.B. Polignando | Algeria |
| 1980 | M.D. Rumbaugh | N W. America |
| 1980 | W.L. Graves | North Africa |
| 1981 | J.H. Elgin; W.F. Lehman, M.D. Rumbaugh | Turkey; Bolivia, Peru, Ecuador |
| 1982 | M.D. Rumbaugh |
Morocco,USSR |
| 1983 | M.D. Rumbaugh | Morocco |
| 1984 | M. D. Rumbaugh and N.L.Taylor |
Romania |
| 1986 | D.A. Johnson |
Pakistan, Nepal |
| 1987 | W. Kaiser | Spain |
| 1988 | K. Asay; N.L. Taylor |
USSR; Yugoslavia |
| 1989 | F.J. Muehlbauer, W.J. Kaiser and C.R. Sperling | Syria and Turkey |
| 1989 | D. Dewey and K. B. Jensen |
Novosibirsk and Altai Mountains |
| 1989 | P. Beuselink, W. Graves, and J. Kirkbride Jr. | Morocco |
| 1990 | K.F.M. Reed and B.S. Dear | USSR |
| 1991 | D. A. Johnson and M.D. Rumbaugh | Xinjiniang, China |
| 1991 | L. Teuber, D. Mowry , T.A. Campbell |
Mongolia |
| 1993 | G.A. Pederson andK.H. Quesenberry | S. Bulgaria |
| 1993 | D.A. Johnson and K.B. Jensen |
N. Xinjiang, China |
| 1993 | G.R.Smith and C.Hoveland | Georgia |
| 1994 | D. A. Johnson and D. P. Sheehy |
Mongolia |
| 1994 | W.L.Graves and P.J.Cunningham | N. Africa |
| 1995 | A.A. Afonin, M.D. Rumbaugh, W. Graves |
Caucasus Mountains, S. Russia |
| 1996 | T. Lumpkin |
Kazakhstan, Kirgizia, and Uzbekistan |
| 1996 | W.J. Kaiser and R. Hannan | Bulgaria |
| 1996 | D.A.Johnson and D.P.Sheehy |
China |
| 1997 | D. A. Johnson, L.K. Holzworth,T.A.Campbell, J.D. Berdahl | China |
| 1998 | D. A. Johnson, D. P. Sheehy, M.E. Majerus, S.R.Winslow |
Mongolia |
| 1999 | R.C. Johnson ,H. Bockelman | Ukraine |
In the early part of the century, alfalfa germplasm was obtained and selectively
distributed by the USDA Office of Foreign Plant Introduction, in Beltsville,
Maryland. In the 1940's the Regional Plant Introduction Stations were established
to increase, evaluate, and distribute plant introductions. As plant introductions
came into the United States, they were distributed to the four Regional PI Stations
for evaluation. By 1949 the North Central Regional Plant Introduction Station
(NCRPIS) in Ames, Iowa became the main site responsible for the increase, evaluation
and distribution of alfalfa plant introductions. In 1971, the USDA National
Plant Germplasm System was established, with the broader mission of preserving
and encouraging the use of germplasm as a guard against genetic erosion and
crop vulnerability. In 1977, NCRPIS set up a specific cooperative agreement
with University of Nevada, Reno, to increase the alfalfa accessions under caged
conditions. This project was terminated in 1986. In 1987 the alfalfa collection
was transferred from Ames, IA to the Western Regional Plant Introduction Station
(WPIS), in Pullman, WA. A Specific Cooperative Agreement was set up with Washington
State University, Prosser to increase the alfalfa accessions under caged conditions.
Efforts in the last two decades have focused on systematically regenerating
the entire collection using controlled pollination, since prior seed increases
had been contaminated by open pollination. A second important emphasis in the
last two decades has been a germplasm evaluation program directed by the ACGC
and administered by WRPIS. The objective of the ACGC has been to obtain a set
of standard descriptors on newly regenerated material, since previous evaluations
had been conducted on accessions representing open pollinated populations. Each
year evaluation proposals were reviewed and funded based on priorities determined
by the ACGC.
By 1988, the USDA alfalfa CRIS project was funded at $160,000. This supported regeneration efforts at Prosser, WA, in the form of supplies and field labor. It also provided support for the germplasm evaluation program managed by the ACGC. In 1993, an alfalfa curator was hired to manage the collection, which had previously been managed by the Research Agronomist, Dr. R.C. Johnson, at the Western Regional PI Station. Whereas the salaries of R.C. Johnson and R. Peaden had not been supported by the Alfalfa CRIS, the alfalfa curator's salary was. In 1995, the Trifolium (5000 accessions) and Lotus (700 accessions) collections were transferred from the Northeastern Regional Plant Introduction Station in Geneva, New York, to WRPIS, in Pullman. The Alfalfa CRIS was rewritten to include the clover and trefoil collections, but no additional funds accompanied the collection transfer. With a flat budget and expanded responsibilities, funds have focused exclusively on regeneration activities. The ACGC-directed germplasm evaluation program was ended in 1997. One of the final projects of the ACGC evaluation program was to develop core subsets of the annual and perennial species.
A core subset is a small subset of accessions from the entire collection that: 1) contains most of the genetic variability that exists in the overall collection, 2) identifies duplications in the collection, 3) recognizes where there is a lack of germplasm representing a species or geographic location, 4) simplifies evaluation especially for difficult and/or expensive traits, and 5) increases the utilization of the collection. There are currently two (2) core subsets related to Medicago, one each for the perennial and annual Medicago species. In addition, development of a Rhizobium meliloti core collection is currently under development.
The perennial Medicago core subset was developed by Drs. Daniel Basigalup, Don Barnes (USDA/ARS) and Robert Stucker at the University of Minnesota in 1989 through 1991. The core subset was selected from 1105 accessions representing 47 different countries and 4 species (M. sativa including the 6 subspecies (ssp. sativa, ssp. varia, ssp. falcata, ssp. coerulea, ssp. falcata, and ssp. ambigua), M. cancellata, M. platycarpa, and M. ruthenica. The 1105 accessions were assembled into 18 different groups based on the original collection site and their geographic proximity, i.e. one group was accessions collected in Hungary, Greece, Italy and Romania. These accessions were evaluated at Rosemount, Minnesota in 1989 for 21 morphological traits and 8 quality traits. Additional evaluation data on these accessions were obtained from the GRIN system including information on the resistance to 10 diseases and 5 insects, and salt and acid tolerance. Selection of the core was made by using cluster analysis to designate the most diverse accessions within each geographic proximity group. The core subset has 200 accessions (~18% of the evaluated accessions) with representatives from each species and geographic area (Basigalup, et. al., 1995).
The annual Medicago core subset was developed by Drs. Noa Diwan, Gary Bauchan (USDA-ARS, Beltsville, MD) and Marla McIntosh at the University of Maryland, College Park, Maryland from 1990 through 1992. The medic core subset was selected from 1220 accessions representing 34 annual Medicago species. These accessions were evaluated in Beltsville, Maryland in 1990 through 1992 for 15 morphological and agronomic traits (days to flower, days to full pod, biomass production within a species, biomass production among species, plant height, plant width (spread), growth habit, middle leaflet length, middle leaflet width, third internode length, pod production, pod spinyness, number of flowers/raceme, number of pods/raceme, and seed size). All of these data are available in the GRIN system. The selection for the core was made using cluster analysis with the final selection of core accessions within each cluster based on geographic location. The annual medic core subset has 211 accessions (~17% of the evaluated accessions) and 34 annual Medicago species are represented (Diwan, et al., 1994). The core collection was evaluated in six (6) locations across the U. S. (Athens, GA, Beltsville, MD, Ithaca, NY, Logan, UT, St. Paul, MN, and Tucson, AZ) for 11 traits (days to flower, days to full pod stage, full pod stage, growth habit, biomass within species, biomass among species, pod production, pod spinyness, plant height, plant width, and winter hardiness). All of these data are available in the GRIN system.
A comprehensive collection of R. meliloti isolates has been developed
at St. Paul, Minnesota. It consists of a domestic collection (U. S. and Canadian)
that was initiated by W. R. Ellis in 1979. That collection was formed by requesting
alfalfa scientists in nearly all states and provinces to send nodules collected
from up to four of the best adapted cultivars. We requested that whenever possible
nodules from Saranac and Saranac AR be included in each locations' collection.
An international collection was initiated by S. E. Viteri in 1987 using the
same procedures of contacting alfalfa scientists. In addition, collections from
Western Australia (Howieson), Napal and Pakistan (Rumbaugh), Turkey (Elgin),
and the USDA (Beltsville, VanBerkum) were also included in the Minnesota collection.
Each isolate was isolated from an individual nodule, increased in culture, inoculated
onto plants from a broadbased Medicago sativa gene pool (BIC-7), and
then reisolated from an effective nodule. All isolates were further tested to
insure that they were R. meliloti.
The entire collection was evaluated for reaction to 13 antibiotic and 14 bacteriophages.
By considering these evaluations it was possible to eliminate similar isolates
from each original collection location. For example, seventeen isolates from
two Georgia fields were tested for bacteriophage reaction. Only two isolates
had a similar profile of reactions. Similarly the two isolates were reconfirmed
with the antibiotic evaluation. After all duplicate isolates were discarded
the collection consisted of 641 isolates. All of these isolates were deposited
in 1990 in the World Rhizobium Culture Collection in the Soybean and
Alfalfa Research Lab., Beltsville, Maryland, Peter VanBerkum, curator. All of
the isolates have been assigned a USDA accession number and have been placed
in cold storage for long-term preservation.
The technical advances of molecular genetics, and interest in developing more sustainable agricultural systems, coupled with the continued rise in loss of biodiversity, have prompt us to closely examine the current contents of the collection to determine if it meets the needs of a broad array of users and conservation objectives. The Convention on Biological Diversity necessitates that this examination occur within the context that nations have sovereign rights over the genetic resources that originate within their boundaries.
Cultivated, naturalized and wild forms of Medicago sativa sbsp.
sativa
Modern and obsolete cultivars
Landraces
Ecotypes
M. sativa ssp. falcata
The following table indicates the number of alfalfa cultivars maintained in the NPGS collection. These accessions have been verified as registered cultivars that were directly donated by the country that released them.
Table 5. Number of cultivars and country of origin represented in the NPGS collection.
| Country |
Number |
Country |
Number |
|---|---|---|---|
| Afghanistan |
1 |
Jordan |
1 |
| Algeria |
2 |
Kazakhstan |
8 |
| Argentina |
13 |
Kenya |
2 |
| Australia |
12 |
Kyrgyzstan |
3 |
| Azerbaijan |
2 |
Lebanon |
1 |
| Belarus |
3 |
Mexico |
8 |
| Belgium |
1 |
Mongolia |
12 |
| Brazil |
2 |
Morocco |
1 |
| Canada |
9 |
New Zealand |
2 |
| Chile |
2 |
Peru |
6 |
| China |
62 |
Poland |
10 |
| Czech Republic |
1 |
Romania |
9 |
| Czechoslovakia |
7 |
Russian Federation |
51 |
| Denmark |
4 |
South Africa |
4 |
| Ecuador |
1 |
Spain |
15 |
| Estonia |
2 |
Sweden |
7 |
| Finland |
1 |
Tajikistan |
2 |
| France |
50 |
Turkey |
3 |
| Germany |
12 |
Ukraine |
11 |
| Hungary |
6 |
United Kingdom |
6 |
| India |
4 |
United States |
47 |
| Iran |
1 |
Uruguay |
1 |
| Israel |
1 |
Uzbekistan |
11 |
| Italy |
13 |
Yemen |
2 |
| Japan |
7 |
Yugoslavia |
6 |
Should the NPGS collection preserve and distribute alfalfa cultivars that were developed in the United States but no longer commercially available? Currently 47 U.S. cultivars are preserved in the active collection. The National Seed Storage Laboratory (NSSL) preserves seed samples of cultivars (and breeding lines and germplasm) that have undergone Crop Science registration (CSR). There are currently over 350 alfalfa cultivars, breeding lines and germplasms that are stored at NSSL. Most of this material has not been assigned PI numbers nor is considered to be part of the active collection that is maintained and distributed from Washington State. Over 800 cultivars have been registered since the inception of the National Alfalfa Variety Review Board, with 100 being registered each year between 1996 and 2000. Obviously, not all these cultivars are CSR and have seed stored at the NSSL. Policy needs to be established regarding the preservation and distribution of obsolete US alfalfa cultivars in the NPGS.
Given that funding can keep pace with a growing collection, the NPGS active collection should support the preservation of US cultivars that are no longer available through commercial sources. Although alfalfa is the fourth most widely grown crop in the US, NGO interest in conserving heirloom alfalfa varieties is poor to non-existent. Efforts need to be made to secure representatives of older varieties before seed stocks deteriorate. Materials registered with Crop Science should enter the active collection when the developer no longer distributes seed or it is not available from commercial sources. As non-CSR material is commercially retired, developers are encouraged to enter the obsolete cultivar into the NPGS collection. When PVP expires, cultivars should become part of the active collection, as PVP suggests the material is of exceptional value. At current levels of funding, a policy that preserves US cultivars as they are retired from commercial production cannot be implemented.
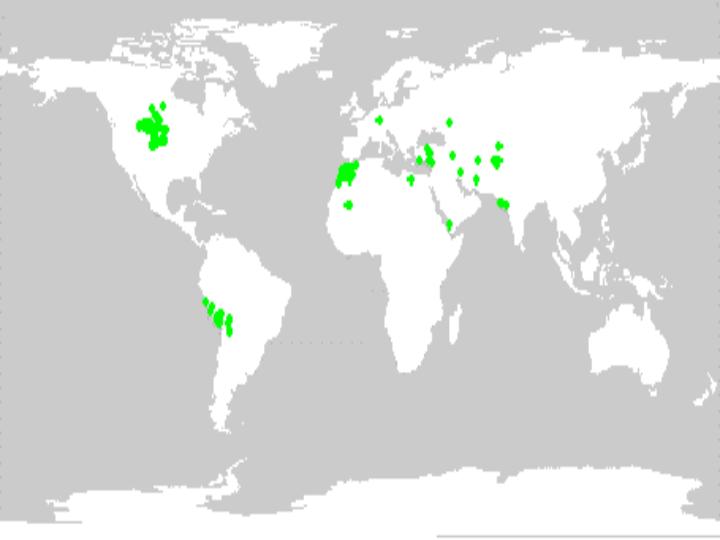
As of 1998, the NPGS collection contained about 600 alfalfa accessions classified as landrace material, and 300 accessions that were classified as cultivated (i.e. data insufficient to make a determination if landrace or cultivar). Figures 6 and 7 indicate the geographic distribution of accessions that had sufficient locality information to obtain map coordinates of collection sites.
Figure 6. Distribution of landrace germplasm held in the
NPGS collection.
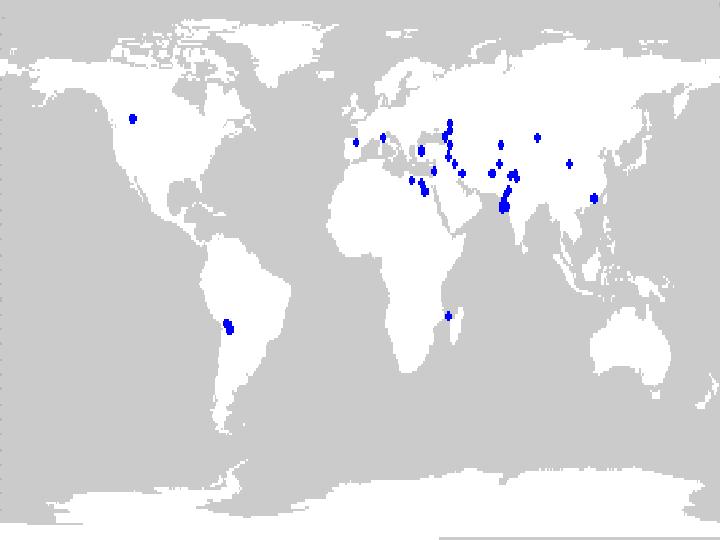
Figure 7. Distribution of cultivated germplasm held in the US collection.
Recommendations
U.S users can make use of collections held in other institutes. Many of the
European nations contain extensive collections of local landraces and primitive
cultivars. For example, the European perennial Medicago database contains
passport data on 2888 accessions of 32 taxa, including 505 land race accessions
stored in 23 European genebanks http://www.cgiar.org/ecpgr/platform/Crops/Medicago.htm)
.
Priority geographic areas recognized by the ACGC in 1988, and not yet collected
include the Himalayan region of India, south and northeast regions of China,
Sudan, Egypt and the Arabian Peninsula. Other geographic areas with limited
representation of landrace materials include South Africa, Northern Kazakhstan,
and adjacent Central Asian countries (i.e. Kyrgistan, Uzbekistan, Afghanistan),
Iran and Iraq.
Past CGC reports have placed emphasis on cultivated alfalfa that is adapted to acid soils and saline soils. There is increasing interest in developing alfalfa with increased water use efficiency, so drought tolerant germplasm would also be valuable to collect
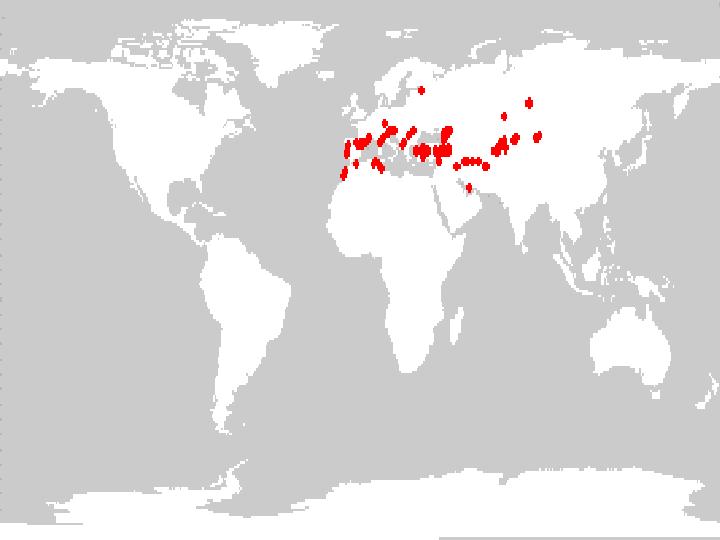
As of 1998, the NPGS collection contained 369 accessions of alfalfa (Medicago sativa subsp. sativa) classified as wild germplasm. Figure 8 indicates the geographic distribution of accessions that had sufficient locality information to obtain map coordinates of collection sites.
Figure 8. Distribution of wild Medicago sativa subsp. sativa germplasm held in the NPGS collection.
Reflecting historic objectives of plant introduction, crop improvement and
preservation of cultivated germplasm, 64 % of the collection consists of alfalfa
accessions (Medicago sativa subsp. sativa). Of the approximately
3000 alfalfa accessions, 73 % represent cultivars, landrace or breeding material.
Wild relatives of cultivated alfalfa remain the most significant gap in the
NPGS collection. The recent success of transferring the glandular hair trait
from close relatives into alfalfa, providing resistance to potato leaf hopper
has been recognized as a major accomplishment in alfalfa breeding. Additionally,
there are substantial breeding efforts engaged in introgressing M. sativa
subsp. falcata germplasm, into alfalfa. In a survey of 63 ex situ
germplasm collections, related species were significantly under-represented
in other collections as well. Acquisition resources need to be focused on this
group of germplasm to provide valuable germplasm for breeders, and ensure the
conservation of these valuable resources.
Table 6. Number of accessions representing wild relatives (primary gene pool) of alfalfa in the NPGS collection.
| Category | ssp. glomerata/ glutinosa |
M. prostrata |
ssp. falcata |
nothossp. varia |
|---|---|---|---|---|
| Breeding |
0
|
0
|
11
|
0
|
| Cultivar |
0
|
0
|
21
|
106
|
| Cultivated |
2
|
0
|
50
|
14
|
| Landrace |
0
|
0
|
20
|
63
|
| Uncertain |
0
|
0
|
18
|
17
|
| Wild |
5
|
13
|
276
|
76
|
| Totals |
7
|
13
|
396
|
276
|
Fill the following geographic gaps in US collection through collection and donations. Species are listed in priority order:
1. Medicago sativa subsp. glomerata (2x=16), subsp. glutinosa
(2x=32) are grouped together since chromosome counts have not been obtained
to distinguish among the two subspecies: Southern Europe, Caucasus region, Algeria,
Tunisia
2. Medicago prostrata: Albania, eastern Austria, Greece, Czechoslovakia,
Hungary, Romania, Yugoslavia
3. M. sativa subsp. falcata: Northern Libya, Eastern Morocco,
Lebanon, Syria, Turkey, Central Asia (Turkmenistan, Afghanistan, Uzbekistan,
Tajikistan, northern Kazakhstan), Northeastern China, Northwest India, Northern
Pakistan, Southern Caucasus region, Ukraine, Northeastern Europe (Belarus, Lithuania,
Latvia, Estonia)
4. M. sativa nothossp varia: Northwest Iran, Syria, Northern,
Western, and Central Europe
Only 1.5% of the total U.S. Medicago collection are species within the secondary gene pool. Representation of the secondary gene pool is also limited in other ex situ collections around the world.
Fill the following gaps in US collection through collection and donations. Species are listed in priority order:
1. M. saxatilis: All areas of species distribution (see Appendix 1)
2. M. papillosa: Northern Armenia, Southern Georgia, Turkey
3. M. sativa nssp. tunetana: All areas of species distribution
(see Appendix 1)
4. M. sativa ssp. caerulea: Crimea, Central Asia
With the exception of M. marina, species in the secondary gene pool are limited in the NPGS collection, and other ex situ collections around the world.
Fill the following gaps in US collection through collection and donations. Species are listed in priority order:
1. M. dzhawakhetica: Caucasus Mountains, Russia
2. M. daghestanica: Caucasus Mountains, Dagesta
3. M. rupestris: Lower Crimea Mountains, Ukraine
4. M. hybrida: Southern France
5. M. rhodopea: Southern Bulgaria
6. M. pironae: Alps of northeastern Italy
7. M. cancellata: Russia (North Caucasus, Caspian region)
Cultivated Medics
1. Most cultivated varieties of annual medic have been developed in Australia. Although they can be used in the southern latitudes of United States (notably California) they lack cold tolerance to more northern latitudes. Acquisition of accessions that are adapted to the northern boundaries of the species distribution may help us develop annual medic cultivars that can be used as cover crops or in ley farming in Washington, Montana and Wyoming.
2. The following cultivated species has been classified by Prosperi et al. as being vulnerable due to lack of representation in ex situ collections: M. arborea, M. italica, M. littoralis, M. rugosa, M. scutellata. Priority should focus on obtaining complete ecogeographic coverage of these taxa.
Non-cultivated Medicago species
Perennial species
Seventy five percent of the perennial Medicago species received a conservation status of "A" by Prosperi et al. (A=endangered species, endemic or rare, very few accessions available in gene banks). With the exception of M. ruthenica, M. arborea and M. marina, the NPGS has few representatives of the perennial wild species.
Recommendations:
Efforts should be made to collaborate with other PGR institutes to conserve these resources in gene bank collections and in natural reserves.Annual species
Representation of annual Medicago species is relatively strong in the NPGS collection since the inclusion of the Lesin's collection from Canada, in 1984. The collection contains about 4000 accessions of 35 species. Although most of these accessions have been increased, many still have not been assigned PI numbers. Documentation of the Lesin's accessions is sparse, and for many accessions, very few seeds were obtained.
Recommendations:
• Efforts should focus on determining if all accessions of the Lesin's collection should receive PI numbers. We need to collaborate with other PGR institutes to conserve those annual species that have potential use, or are vulnerable to genetic erosion.
• Efforts should be made to collaborate with other PGR institutes to conserve these resources in gene bank collections and in natural reserves.
Since 1981 the perennial Medicago collection has been evaluated for 14 diseases, six (6) insects, chromosome number, seven (7) feed quality traits, 25 agronomic traits, and five (5) stress tolerant traits. All of the evaluations were conducted using standardized tests when they were evaluated. Scientists in the federal government, universities and private industry conducted the tests and provided the data which was imported into GRIN. Appendix 2 is a listing of the evaluations which have been conducted. They indicate the percentage of accessions for each species that have been evaluated for a specific descriptor. In some cases accessions have been evaluated multiple times for the same descriptor. The percentages reflect only a single evaluation event per accession.
The majority of annual medic accessions have data that characterize production, phenology and growth (Appendix 3). These data were acquired as accessions and were regenerated in Riverside, CA from 1988 to 1999 and as the annual medic core collection was developed in Beltsville, MD and evaluated in six (6) locations (Tucson, AZ; Athens, GA; Beltsville, MD; St. Paul, MN; Ithaca, NY and Logan, UT) across the U.S.
• Data on only one disease descriptor has been gathered, and for only the core collection, thus, additional diseases need to be screened.
• Some morphological descriptors have been taken, however, the majority of the accessions in the collection do not have even basic morphological descriptors. Thus a complete set of morphological descriptors needs to be obtained.
Because the current PI collection of perennial Medicago spp. is large and mostly in "raw" form (i.e., without many of the traits necessary in current varieties), it is not practical for commercial breeders to use for developing alfalfa varieties with improved quantitative traits (forage and seed yield, quality, etc.). Current market competition demands rapid development of "new and improved" varieties; therefore, most breeders have insufficient time and resources to improve non-elite germplasm into forms useable for varietal development. Therefore, a national effort to genetically enhance these PI's is necessary for their utilization in programs to develop varieties.
Simply inherited traits from unadapted PI's may be incorporated into adapted
backgrounds by backcrossing or by biotechnological methods available in the
future. For quantitative traits, however, early use of PI's probably will be
in the form of adapted or nearly adapted germplasm pools (GPP). These pools
should have at least moderate levels of the economic traits available in current
varieties, while containing large variability for traits that may be useful
to improve alfalfa in the future. Alfalfa breeders need GPP's as sources to
tap both for "new" genes conditioning traits currently recognized
as important and for traits that may be needed in the future. These pools also
would provide material for research in alternate uses of alfalfa, such as sustainable
agriculture.
An enhancement proposal was submitted to the ACGC in 1990 which outlined methods
to develop alfalfa GPP's that should be useful to breeders in the USA. The proposal
was developed by a subcommittee within the ACAC and it serves as an initial
focal point from which the ACGC can plan for developing national and regional
GPP's. The main objectives of the proposal are:
1. Develop a national GPP to serve as a source of new recombinants among the total diversity of germplasm available. This pool potentially would provide recombinants not obtained in the more genetically restricted regional pools described below.
2. Develop regional GPP's that are adapted to each of four regions in the USA. Under the typical management practices for alfalfa in a particular region, the regional pools should be agronomically more acceptable than the national GPP. They also should have at least moderate levels of the traits available in varieties currently used in that region, plus genes that may be used to improve cultivated alfalfa.
• The proposed enhancement program for the development of national and regional germplasm pools should be enacted. Appendix 4 provides details of the enhancement program.
• An ARS alfalfa research geneticist position was established in 1999 at Prosser, WA. with an emphasis to carry out additional germplasm enhancement work. Enhancement work by this position should continue to be encouraged and supported.
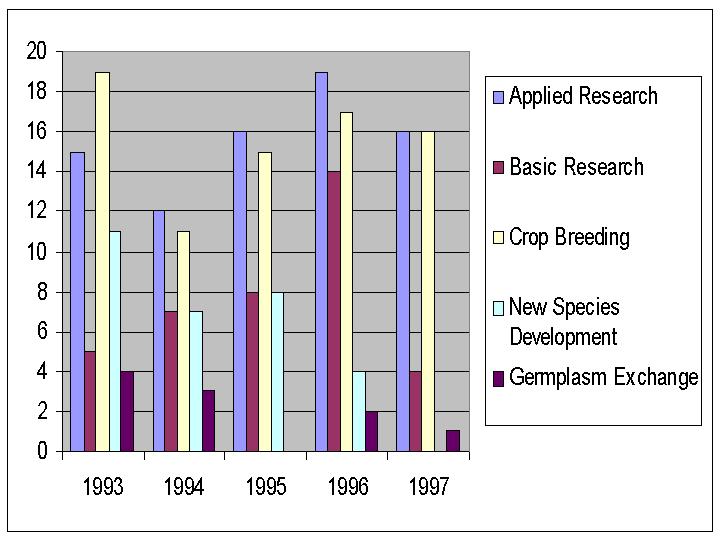
A total of 12,800 seed packets were distributed for 229 requests from 1993 through 1997. Scientists from U.S. universities were responsible for almost half of the seed requests. The remaining requests for perennial species were split about equal between USDA-ARS scientists, private breeding companies and foreign scientists. For annual species, private company requests accounted for only 4%. The majority of requested germplasm was intended for applied and basic research projects. A substantial amount of germplasm was requested for screening by crop breeding programs.
When collection use was examined by taxa, and size of collection factored out, use of germplasm representing the secondary and tertiary alfalfa gene pools has been higher on average, than use of the alfalfa collection in the last 5 years. However, 5400 seed packets were distributed from the M. sativa ssp sativa collection. Requests for non-cultivated annual species has been greater than for the perennial species and is probably due to requests for the annual medic core collection .
Five (5) % of the orders for perennial species were for the alfalfa core collection, 13 % of orders for annual species were for the core subset.
Figure 9. Number of requests grouped by intended seed use as reported by requestor from 1993 to 1997.
The Medicago collection is housed and distributed from the USDA, ARS Western Regional Plant Introduction Station located in Pullman, WA. Original seed packets and regeneration seed lots are kept at 38-40 Co with 28 % relative humidity. Seed lots that are distributed are kept at the same conditions. Seed counts have been obtained on all packets except for original seed. Germination testing has been carried out for seed lots increased in 1996, 1997 and 1998. Original seed has not had germination tests, nor have baseline germinations been obtained on increase seed lots. Approximately 84 % of the accessions in the collection have a safety backup sample stored at NSSL. Accessions not backed up are those with very few seeds, or are currently queued for regeneration.
From 1979 to 1997, the majority of perennial accessions were regenerated using original or the oldest existing seed lot, under isolation in cages using up to 200 plants per population (this number varies with amount of seed available; number of plants harvested is placed in GRIN as a historic record). From 1987 to 1998, most of the annual medic species were increased at Riverside, California. Accessions in the Medicago collection that remain to be increased include those species that are difficult to regenerate, and accessions with very low seed counts or low seed viability. These accessions require special handling to ensure they are not lost during the regeneration process. Approximately 500 accessions fall into this category.
Original seed needs to be inventoried with seed counts. Germination data on original seed lots needs to be obtained on all accessions with sufficient seed. Original and regeneration seed lots need to be stored at –15 to –20 C.
Germination data needs to be obtained on increased material to establish a
baseline to monitor seed viability and to provide a check to verify that regeneration
procedures are high quality.
Research needs to be carried out to verify if our standard plant population
size during regeneration is adequate for limiting genetic drift, and if genetic
change in accessions is minimized during the standard regeneration process.
The genus Medicago according to Small and Jomphe, (1989), with geographic
distribution, number of accessions per species1 in the NPGS collection,
and conservation status of each species as defined by Properi et al.
(1996) .
| Section |
Subsection |
Species |
Subspecies |
Geographic Distribution |
NPGS Accessions |
Status1 |
|---|---|---|---|---|---|---|
| Dendrotelis | M. arborea L. S. | Europe, Turkey | 38 | B-Cult | ||
| Dendrotelis | M. strasseri Greuter, Matthas & Risse | Crete | A | |||
| Medicago | M. sativa | ssp. caerulea Schmalh. | E. Turkey, Iran, Former USSR |
77 | B | |
| Medicago | M. sativa | ssp. sativa (L.) L & L. | Possible native range-Middle East, C Asia, now widely distributed |
3042 |
C-CULT | |
| Medicago | M. sativa | ssp. glomerata | Balbis. S Europe, N Africa, Caucasus | 7 | A | |
| Medicago | M. sativa | ssp. falcata (L.) Arcangeli | S Europe, Former USSR | B-CULT | ||
| Medicago | M. sativa | var falcata Archang. | N Eurasia | 419 | - | |
| Medicago | M. sativa | var viscosa Posp. | S Europe, N Africa, Caucasus | 14 | - | |
| Medicago | M. sativa | ssp. x varia Martin | Europe, Iran, Syria, Turkey, Caucasus | 287 | - | |
| Medicago | M. sativa | ssp. x tunetana Murbeck | S Europe, N Africa, Caucasus | 6 | - | |
| Medicago | M. papillosa Boiss. | Turkey, S Russia | 8 | A | ||
| Medicago | M. papillosa | ssp. macrocarpa (Boiss) Urban | Turkey | 0 | - | |
| Medicago | M. papillosa | ssp. papillosa Bioss. | Turkey, S Russia | 1 | - | |
| Medicago | M. prostrata Jacq. | Albania, E Europe, Italy | 13 | A | ||
| Medicago |
M. rupestris M. Bieb. |
Crimea, W Caucasus | 0 | A | ||
| Medicago | M. cancellata M. Bieb. | European Russia | 7 | A | ||
| Medicago |
M. rhodopea Velen. |
Bulgaria | 2 | A | ||
| Medicago | M. saxatilis M. Bieb. | Crimea | 1 | A | ||
| Medicago | M. daghestanica Rupr. Ex Boiss | Dagestan | 0 | A | ||
| Medicago | M. pironae Vis. | Italy | 4 | A | ||
| Medicago |
M. hybrida (Pourret) Trautv. |
France, Spain | 1 | A | ||
| Medicago | M. suffruticosa Raymond ex DC. | France, Spain, Morocco | 17 | B | ||
| Medicago | M. marina L. | S Europe, N Africa, Middle East, Crimea | 20 | B | ||
|
Carstiensae |
M. carstiensis Jacq. | E Europe | 1 | A | ||
| Spirocarpos | Pachyspirae | M. soleirolii Duby | France, Italy, Algeria, Tunisia | 11 | A | |
| Spirocarpos | Pachyspirae |
M. italica (Miller) Fiori |
Mediterranean Basin | 87 | B-CULT | |
| Spirocarpos | Pachyspirae | M. littoralis Rohde ex Lois | Mediterranean Basin, E Europe, Caucasus | 173 | V | |
| Spirocarpos | Pachyspirae |
M. truncatula Gaertner |
Mediterranean Basin, E Europe, Caucasus |
320 |
C-CULT |
|
| Spirocarpos | Pachyspirae | M. doliata Carmign. | Italy, Spain, Algeria, Morocco | 104 | C | |
| Spirocarpos | Pachyspirae |
M. turbinata (L.) All. |
Mediterranean Basin |
86 |
B |
|
| Spirocarpos | Pachyspirae | M. rigidula (L.) All. | Mediterranean Basin, E Europe, Caucasus, Central Asia | 148 | C-CULT | |
| Spirocarpos | Pachyspirae |
M. rigiduloides Small. |
E Mediterranean, Middle East |
192 |
||
| Spirocarpos | Pachyspirae |
M. constricta Durieu |
E Mediterranean Basin, Iran, Iraq |
52 |
B |
|
| Spirocarpos | Pachyspirae |
M. lesinsii E. Small |
Mediterranean Basin |
9 |
B-CULT |
|
| Spirocarpos | Pachyspirae |
M. murex Willd. |
Mediterranean Basin |
69 |
B-CULT |
|
| Spirocarpos | Pachyspirae | M. rugosa Desr. | Mediterranean Basin | 43 | B | |
| Spirocarpos | Pachyspirae |
M. scutellata (L.) Miller |
Mediterranean Basin, Southern Ukraine, Crimea |
61 | B-CULT | |
| Spirocarpos | Pachyspirae |
M. blancheana Boiss. |
E Mediterranean |
18 |
B |
|
| Spirocarpos | Rotatae | M. rotata Boiss. | Cyprus, Iraq, Israel, Jordan, Lebanon, Syria, Turkey |
21 |
B | |
| Spirocarpos | Rotatae |
M. noeana Boiss. |
Iraq, Turkey |
19 |
A |
|
| Spirocarpos | M. shepardii Post. | Ex Boiss. Turkey | 4 | A | ||
| Spirocarpos | Intertextae | M. intertexta (L.) Miller | W Mediterranean Basin | 17 | B | |
| Spirocarpos | Intertextae |
M. ciliaris (L.) Krocker |
Mediterranean Basin, Iraq |
64 |
C |
|
| Spirocarpos | Intertextae | M. muricoleptis Tineo | France, Italy | 8 | A | |
| Spirocarpos | Intertextae | M. granadensis Willd. | Egypt, Israel, Syria, Turkey | 13 | B | |
| Spirocarpos | Leptospirae | M. sauvagei Negre | Morocco | 5 | A | |
| Spirocarpos | Leptospirae |
M. laciniata (L.) Miller |
N Africa, Arabian peninsula, India, Pakistan, Afghanistan |
136 |
B | |
| Spirocarpos | Leptospirae | M. minima (L.) Bartal. | Europe, N Africa, India., Russia | 299 | C | |
| Spirocarpos | Leptospirae | M. praecox DC. | S and E Europe, Cyperus, Turkey, Crimea | 23 | B | |
| Spirocarpos | Leptospirae |
M. coronata (L.) Bartal. |
E Mediterranean Basin, Iran, Iraq |
24 |
B |
|
| Spirocarpos | Leptospirae |
M. polymorpha L. |
Europe, N Africa, Middle East, Crimea, Caucasus, Central Asia |
685 |
C-CULT | |
| Spirocarpos | Leptospirae |
M. laxispira |
Heyn. Iraq | 0 | A | |
| Spirocarpos | Leptospirae | M. arabica (L.) Hudson | Europe, N Africa, Middle East, Crimea, Caucasus | 79 | C | |
| Spirocarpos | Leptospirae |
M. tenoreana Ser. |
France, Italy, Yugoslavia |
6 |
A |
|
| Spirocarpos | Leptospirae | M. disciformis DC. |
Mediterranean Basin | 50 | B |
|
| Spirocarpos | Leptospirae | M. lanigera Winkler & B. Fedtsch. | Afghanistan, Turkmenistan, Tajikistan |
1 | A | |
|
Geocarpa E. Small |
Leptospirae | M. hypogaea E. Small | SE Mediterranean | 0 | A | |
| Lupularia |
M. lupulina L. |
Europe, N Africa, Middle East, Asia |
209 |
C-Cult |
||
| Lupularia | M. secundiflora Durieu | S Europe, N Africa | 2 | B | ||
| Heynianae | M. heyniana Greuter | Greece | 2 | A | ||
| Orbiculares |
M. orbicularis (L.) Bartal. | Mediterranean basin, Middle East, Central Asia | 322 | C | ||
| Hymenocarpos |
M. radiata L. | Middle East, Russia, Central Asia | 11 | C | ||
| Platycarpae |
M. plicata (Boiss) | Turkey | 0 | UNK | ||
| Platycarpae | M. platycarpa L. | Trautv. China, Mongolia, Central Asia | 15 | A | ||
| Platycarpae |
M. ruthenica L. Trautv. |
China, Korea, Mongolia, Russia |
122 |
A |
||
| Platycarpae | M. popovii Sirj. | Central Asia | 1 | A | ||
| Platycarpae | M. archiducis-nicolai Sirjaev | Central China, NE Tibet | 0 | UNK | ||
| Platycarpae | M. edeworthii Sirjaev | Himalayas | 3 | A | ||
| Platycarpae | M. cretacea M. Bieb. | Russia, Former USSR | 2 | A | ||
| Platycarpae | M. ovalis (Boiss.) Sirjaev | S Spain, Morocco | 0 | UNK | ||
| Lunatae |
M. rostrata (Boiss. & Bal.) | Turkey | 0 | ? | ||
| Lunatae |
M. biflora (Griseb.) E Small |
Turkey, Iran, S Trans-Caucasus | 1 | ? | ||
| Lunatae | M. brachycarpa | Turkey, Lebanon, Iraq, Transcaucasia | 5 | ? | ||
| Lunatae | M. huberi E. Small | SW Anatolia | 0 | ? | ||
| Buceras | Erectae | M. astroites (Fisch. & Mey.) | E Mediterranean | 0 | ? | |
| Buceras | Erectae | M. halophila (Boiss) E Small | S Anatolia | 0 | ? | |
| Buceras | Erectae | M. arenicola (Huber-Mor) | Turkey | 0 | ? | |
| Buceras | Erectae | M. heldreichii (Boiss.) E. Small | Turkey | 0 | ? | |
| Buceras | Erectae | M. phrygia (Boiss.& Bal.) | Turkey, Syria, Iraq, Iran | 0 | ? | |
| Buceras | Erectae | M. fischeriana (Ser.) Trautv. | Turkey, Iraq, Iran, European USSR | 5 | ? | |
| Buceras | Erectae | M persica (Boiss.) E. Small | Iran, Iraq | 0 | ? | |
| Buceras | Erectae | M. medicaginoides (Retz.) E. Small | SE Europe, SW USSR, Turkey, Iran, S and C Asia | 1 | ? | |
| Buceras | Erectae | M. rigida (Boiss. & Bal.) | S Anatolia | 0 | ? | |
| Buceras | Erectae | M. crassipes (Boiss) E. Small | Anatolia. Lebanon, N Iraq, W Iran | 1 | ? | |
| Buceras | Erectae | M. pamphylica (Huber-Mor. & Sirjaev) E. Small | S Anatolia | 0 | ? | |
| Buceras | Erectae | M. carica (Huber-Mor. & Sirjaev) E. Small | S W Anatolia | 0 | ? | |
| Buceras | Erectae | M. monantha (C.V. Meyer) Trautv. | Middle East, S and C Asia | 38 | ? | |
| Buceras | Erectae |
M. orthoceras (Kar. &Kir.) Trautv. |
Middle East, S and C Asia |
0 |
? |
|
| Buceras | Erectae |
M. polyceratia (L.) Trautv. |
W Mediterranean |
10 |
? |
|
|
|
Deflexae |
M. retrorsa (Boiss.) E. Small |
Afghanistan |
0 |
? |
|
| Buceras |
Reflexae | M. monspeliaca (L.) Trautz. | W Europe, N Africa, E Mediterranean, Jordan, Iraq, Iran, USSR, C Asia | 12 | ? | |
| Buceras |
Isthmocarpae |
M. rhytidiocarpa (Boiss. & Bal.) E. Small | S Anatolia | 0 | ? | |
| Buceras | Isthmocarpae |
M. isthmocarpa (Boiss. & Bal.) E. Small |
C Anatolia |
0 |
? |
1Conservation Status : A = Endangered species, endemic or rare,
very few available accessions in gene banks; B = Species with threatened diversity
due to a rapid modification of its ecosystem or due to its localization in regions
subject to an important climate change, few available accessions; C = Species
widespread, large number of available accessions, Cult = Genetically improved
for agriculture; ? = unknown or undetermined (Prosperi et al. 1996)
| Species | Cytology Disease | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CHR | ANTH | APHA | BACT | DOWN | FUSW | LSPT | LEPT | PHYR | RHIZ | SCLE | SPRB | STEMLF | STEM | VERT | |
| arborea | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| cancellata |
0 |
43 |
43 | 57 | 86 | 57 | 29 | 43 | 14 | 43 | 100 | 43 | 43 | 43 | 86 |
| carstiensis | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| hybrid |
0 |
33 |
33 |
11 |
11 |
11 |
11 |
17 |
17 |
22 |
39 |
17 |
22 |
17 |
28 |
| papillosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 0 |
| subsp. papillosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| pironae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 50 | 0 | 0 | 0 | 0 |
| platycarpos | 0 | 7 | 7 | 13 | 7 | 13 | 0 | 0 | 7 | 7 | 40 | 7 | 7 | 7 | 13 |
| polymorpha | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| prostrata | 15 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 38 | 0 | 0 | 0 | 0 |
| ruthenica | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 16 | 1 | 0 | 1 | 1 |
| sativa nssp. tunetana | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 33 | 0 | 0 | 0 | 0 |
| sativa nssp. varia | 0 | 24 | 54 | 40 | 37 | 40 | 14 | 22 | 29 | 31 | 41 | 22 | 45 | 22 | 67 |
| sativa subsp. caerulea | 38 | 36 | 47 | 52 | 51 | 52 | 31 | 32 | 22 | 9 | 23 | 32 | 36 | 32 | 57 |
| sativa subsp. falcata | 63 | 15 | 18 | 19 | 13 | 19 | 9 | 11 | 8 | 11 | 10 | 11 | 14 | 11 | 23 |
| sativa subsp. glomerata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 43 | 0 | 0 | 0 | 14 |
| sativa subsp. sativa | 1 | 35 | 35 | 32 | 33 | 32 | 28 | 29 | 26 | 21 | 26 | 29 | 32 | 29 | 37 |
| sativa var. viscosa | 15 | 8 | 0 | 8 | 8 | 8 | 8 | 8 | 0 | 0 | 46 | 8 | 0 | 8 | 23 |
| saxatilis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| scutellata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 |
| suffruticosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 15 | 0 | 0 | 0 | 0 |
CHR= chromosome number; ANTH= Anthracnose; APHA= Aphanomyces; BACT= Bacterial wilt ; DOWN= Downey mildew; FUSW= Fusarium wilt; LSPT = Common leaf spot; LEPT= Lepto spot; PHYR= Phytophthera root rot; RHIZ= Rhizoctonia; SCLE= Scleorotinia; SPRB= Spring black stem; STEMLF= Stemphylium leaf spot; STEM= Summer black stem; VERT= Verticilium wilt
1 Tables indicate the percentage of accessions for each species that have been evaluated for a specific descriptor. In some cases accessions have been evaluated multiple times for the same descriptor. The percentages reflect only a single evaluation event per accession.
| Species | Insect | Chemical | |||||||
|---|---|---|---|---|---|---|---|---|---|
| BLUE | LYGUS | PEAA | POTLF | ROOT | SPOT | CLEAF | CSTEM | PBYPASS | |
| arborea | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| cancellata | 43 | 0 | 29 | 100 | 86 | 57 | 43 | 43 | 29 |
| carstiensis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| hybrid | 11 | 6 | 11 | 22 | 22 | 22 | 22 | 22 | 17 |
| papillosa | 0 | 0 | 0 | 13 | 25 | 0 | 0 | 0 | 0 |
| subsp. papillosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| pironae | 0 | 0 | 0 | 50 | 50 | 0 | 0 | 0 | 0 |
| platycarpos |
0 |
7 |
0 |
7 |
13 |
7 |
7 |
7 |
0 |
| polymorpha | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| prostrata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| ruthenica | 0 | 1 | 0 | 2 | 2 | 1 | 0 | 0 | 1 |
| sativa nssp. tunetana | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sativa nssp. varia | 33 | 7 | 33 | 45 | 43 | 38 | 45 | 45 | 8 |
| sativa subsp. caerulea | 52 | 0 | 52 | 52 | 53 | 52 | 36 | 36 | 6 |
| sativa subsp. falcata | 12 | 0 | 12 | 24 | 24 | 14 | 14 | 14 | 3 |
| sativa subsp. glomerata | 0 | 0 | 0 | 14 | 14 | 0 | 0 | 0 | 14 |
| sativa subsp. sativa | 28 | 5 | 23 | 37 | 36 | 32 | 33 | 32 | 6 |
| sativa var. viscosa | 8 | 0 | 8 | 31 | 31 | 8 | 0 | 0 | 23 |
| saxatilis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| scutellata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| suffruticosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 |
BLUE=Blue alfalfa aphid; LYGUS=Lygus bug ; PEAA= Pea alfalfa aphid; POTLF=Potato leaf hopper; ROOT= Clover root curculio; SPOT= Spotted alfalfa aphid; CLEAF= Crude protein in leaf ; CSTEM= Crude protein in stem; PBYPASS= By pass protein
| Species | Growth | |||||||
|---|---|---|---|---|---|---|---|---|
| CSTD |
CWID |
FALL |
GHAB |
RECO | RTD2 | SDLGV | TAPDIA | |
| arborea | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| cancellata | 43 | 43 | 43 | 29 | 29 | 43 | 43 | 43 |
| carstiensis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| hybrid | 22 | 22 | 22 | 11 | 11 | 22 | 22 | 22 |
| papillosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| subsp. papillosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| pironae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| platycarpos | 7 | 7 | 7 | 7 | 7 | 7 | 7 | 7 |
| polymorpha | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| prostrata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| ruthenica | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 |
| sativa nssp. tunetana | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sativa nssp. varia | 45 | 45 | 45 | 24 | 22 | 45 | 45 | 45 |
| sativa subsp. caerulea | 36 | 36 | 36 | 30 | 18 | 36 | 36 | 36 |
| sativa subsp. falcata | 14 | 14 | 14 | 8 | 8 | 14 | 14 | 14 |
| sativa subsp. glomerata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sativa subsp. sativa | 33 | 33 | 33 | 20 | 19 | 33 | 33 | 33 |
| sativa var. viscosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| saxatilis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| scutellata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| suffruticosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
CSTD=Crown stem diameter; CWID Crown width; FALL= Fall growth; GHAB= Growth habit; RECO= Recovery rate; RTD2= Secondary root diamter; SDLGV= Seedling vigor; TAPDIA= Tap root diameter
| Species | Morphology | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CBRAN | CORIG | CBUD | DCRO | DTAP% | DTAPP | FRTM | FLWC | LFSZ | MAT | RNUM2 | RPOS2 | UILEN | |
| arborea | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| cancellata | 43 | 43 | 43 | 43 | 43 | 43 | 43 | 0 | 43 | 43 | 43 | 43 | 14 |
| carstiensis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| hybrid | 22 | 22 | 22 | 22 | 22 | 22 | 22 | 0 | 22 | 22 | 22 | 22 | 6 |
| papillosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| subsp. papillosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| pironae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| platycarpos | 7 | 7 | 7 | 7 | 7 | 7 | 7 | 0 | 7 | 7 | 7 | 7 | 7 |
| polymorpha | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| prostrata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 15 | 0 | 0 | 0 | 0 | 0 |
| ruthenica | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 |
| sativa nssp. tunetana | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sativa nssp. varia | 45 | 45 | 45 | 45 | 45 | 45 | 45 | 0 | 45 | 45 | 45 | 45 | 8 |
| sativa subsp. caerulea | 36 | 36 | 36 | 36 | 36 | 36 | 36 | 0 | 36 | 36 | 36 | 36 | 6 |
| sativa subsp. falcata | 14 | 14 | 14 | 14 | 14 | 14 | 14 | 41 | 14 | 14 | 14 | 14 | 3 |
| sativa subsp. glomerata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sativa subsp. sativa | 33 | 33 | 33 | 32 | 33 | 33 | 33 | 0 | 33 | 32 | 33 | 33 | 6 |
| sativa var. viscosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| saxatilis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| scutellata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| suffruticosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
CBRAN=Crown branching; CORIG= Crown origin; CBUD= Crown budding; DCRO= Depth of crown; DTAP%= Determinate tap root percentage; DTAPP= Determinate tap root position; FRTM= Fiberous root mass; FLWC= Flower color; LFSZ= Leaf size; MAT= Maturity; RNUM2=Secondary root number; RPOS2= Secondary root position; UILEN= Unifoliate internode length
| Species | Production | Stress-Abiotic | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ADFL | ADFS | NDFL | NDFS | SDYLD | SDPROD | SDWGT | SLRAT | ACID | FRDAM | SGERM | SSTRESS | WININJ | |
| arborea | 0 | 0 | 0 | 0 | 0 | 0 | 9 | 0 | 0 | 0 | 0 | 0 | 0 |
| cancellata | 43 | 43 | 43 | 43 | 43 | 100 | 71 | 43 | 43 | 43 | 57 | 29 | 43 |
| carstiensis | 0 | 0 | 0 | 0 | 0 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| hybrid | 22 | 22 | 22 | 22 | 22 | 50 | 61 | 22 | 11 | 22 | 33 | 28 | 22 |
| papillosa | 0 | 0 | 0 | 0 | 0 | 100 | 25 | 0 | 13 | 0 | 0 | 0 | 0 |
| subsp. papillosa | 0 | 0 | 0 | 0 | 0 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| pironae | 0 | 0 | 0 | 0 | 0 | 100 | 50 | 0 | 50 | 0 | 0 | 0 | 0 |
| platycarpos | 7 | 7 | 7 | 7 | 7 | 47 | 27 | 7 | 27 | 7 | 7 | 7 | 7 |
| polymorpha | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| prostrata | 0 | 0 | 0 | 0 | 0 | 92 | 0 | 0 | 23 | 0 | 0 | 0 | 0 |
| ruthenica | 0 | 0 | 0 | 0 | 0 | 90 | 82 | 0 | 6 | 0 | 1 | 1 | 0 |
| sativa nssp. tunetana | 0 | 0 | 0 | 0 | 0 | 50 | 17 | 0 | 33 | 0 | 0 | 0 | 0 |
| sativa nssp. varia | 45 | 45 | 45 | 45 | 45 | 71 | 58 | 45 | 21 | 45 | 60 | 48 | 45 |
| sativa subsp. caerulea | 36 | 36 | 36 | 36 | 36 | 58 | 44 | 36 | 39 | 36 | 48 | 39 | 36 |
| sativa subsp. falcata | 14 | 14 | 14 | 14 | 14 | 45 | 24 | 14 | 10 | 14 | 18 | 16 | 14 |
| sativa subsp. glomerata | 0 | 0 | 0 | 0 | 0 | 86 | 0 | 0 | 29 | 0 | 14 | 0 | 0 |
| sativa subsp. sativa | 33 | 32 | 33 | 32 | 32 | 56 | 69 | 32 | 19 | 33 | 39 | 22 | 33 |
| sativa var. viscosa | 0 | 0 | 0 | 0 | 0 | 77 | 0 | 0 | 31 | 0 | 15 | 0 | 0 |
| saxatilis | 0 | 0 | 0 | 0 | 0 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| scutellata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| suffruticosa | 0 | 0 | 0 | 0 | 0 | 100 | 8 | 0 | 15 | 0 | 0 | 0 | 0 |
ADFL= Acid detergent fiber leaf; ADFS= Acid detergent fiber stem; NDFL= Neutral detergent fiber leaf; NDFS=Neutral detergent fiber stem; SDYLD= Seed yield; SDPROD= Seedling year productivity; SDWGT= 100 seed weight; SLRAT= Stem leaf ratio; ACID= Acid soil tolerance; FRDAM= Frost damage; SGERM= Salt germination; SSTRESS- Tolerance to salt stress; WININJ= Winter injury
| Species | Cytology | Disease | Growth | Morphology | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CHR | ANT | GHAB | WVIG | COIL | DAYSP | HTH | SPRD | SPIN | SCUR | |
| arabica | 0 | 3 | 67 | 86 | 0 | 3 | 3 | 3 | 3 | 0 |
| blancheana |
0 |
39 |
83 |
72 |
0 |
50 | 50 | 50 | 50 | 0 |
| brachycarpa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| ciliaris | 0 | 9 | 77 | 73 | 0 | 8 | 9 | 9 | 8 | 0 |
| constricta | 0 | 6 | 62 | 60 | 0 | 6 | 6 | 6 | 6 | 0 |
| coronata | 0 | 8 | 100 | 100 | 0 | 8 | 8 | 8 | 8 | 0 |
| disciformis | 0 | 8 | 86 | 100 | 0 | 8 | 8 | 8 | 8 | 0 |
| doliata | 0 | 2 | 89 | 88 | 0 | 3 | 3 | 3 | 3 | 0 |
| var. doliata | 0 | 0 | 33 | 33 | 0 | 0 | 0 | 0 | 0 | 0 |
| var. muricata | 0 | 5 | 35 | 33 | 0 | 0 | 0 | 0 | 0 | 0 |
| fischeriana | 0 | 0 | 40 | 40 | 0 | 0 | 0 | 0 | 0 | 0 |
| granadensis | 0 | 31 | 100 | 100 | 0 | 23 | 23 | 23 | 23 | 0 |
| heyniana | 0 | 50 | 50 | 0 | 0 | 50 | 50 | 50 | 50 | 0 |
| hybrid | 0 | 0 | 22 | 22 | 0 | 0 | 0 | 0 | 0 | 0 |
| intertexta | 0 | 35 | 94 | 88 | 0 | 24 | 24 | 24 | 24 | 0 |
| italica | 0 | 10 | 71 | 66 | 0 | 7 | 10 | 8 | 7 | 0 |
| laciniata | 0 | 7 | 81 | 85 | 0 | 7 | 8 | 8 | 7 | 0 |
| lanigera | 0 | 100 | 100 | 100 | 0 | 100 | 100 | 100 | 100 | 0 |
| lesinsii | 0 | 22 | 89 | 89 | 0 | 11 | 11 | 11 | 11 | 0 |
| littoralis | 0 | 4 | 45 | 42 | 0 | 2 | 3 | 2 | 2 | 0 |
| lupulina | 0 | 7 | 48 | 49 | 0 | 8 | 8 | 8 | 8 | 0 |
| medicaginoides | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| minima | 0 | 1 | 71 | 86 | 0 | 1 | 2 | 1 | 1 | 0 |
| monantha | 0 | 0 | 32 | 32 | 0 | 0 | 0 | 0 | 0 | 0 |
| monspeliaca | 0 | 0 | 17 | 17 | 0 | 0 | 0 | 0 | 0 | 0 |
| murex | 1 | 7 | 72 | 86 | 0 | 4 | 7 | 6 | 4 | 0 |
| muricoleptis | 0 | 13 | 88 | 88 | 0 | 0 | 0 | 0 | 0 | 0 |
| noeana | 0 | 11 | 100 | 100 | 0 | 11 | 11 | 11 | 11 | 0 |
| orbicularis | 0 | 2 | 65 | 65 | 0 | 2 | 3 | 3 | 2 | 0 |
| polyceratia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| polymorpha | 0 | 3 | 79 | 78 | 0 | 1 | 1 | 1 | 1 | 0 |
| var. brevispina | 0 | 17 | 80 | 89 | 0 | 20 | 20 | 20 | 20 | 0 |
| var. polymorpha | 0 | 11 | 68 | 78 | 0 | 14 | 14 | 14 | 14 | 0 |
| var. vulgaris | 0 | 0 | 69 | 69 | 0 | 8 | 8 | 8 | 8 | 0 |
| popovii | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| praecox | 0 | 9 | 70 | 83 | 0 | 9 | 9 | 9 | 9 | 0 |
| radiata | 0 | 36 | 55 | 18 | 0 | 36 | 36 | 36 | 36 | 0 |
| rhodopea | 0 | 0 | 50 | 50 | 0 | 0 | 0 | 0 | 0 | 0 |
| rigidula | 0 | 2 | 53 | 82 | 76 | 3 | 3 | 3 | 3 | 76 |
| rigiduloides | 0 | 2 | 59 | 76 | 100 | 1 | 2 | 1 | 1 | 100 |
| rotata | 0 | 29 | 86 | 86 | 0 | 29 | 29 | 29 | 29 | 0 |
| rugosa | 0 | 26 | 88 | 86 | 0 | 19 | 26 | 26 | 19 | 0 |
| sauvagei | 0 | 40 | 100 | 100 | 0 | 40 | 40 | 40 | 40 | 0 |
| scutellata | 0 | 28 | 80 | 80 | 0 | 30 | 31 | 31 | 30 | 0 |
| secundiflora | 0 | 0 | 100 | 100 | 0 | 0 | 0 | 0 | 0 | 0 |
| shepardii | 0 | 0 | 100 | 100 | 0 | 0 | 0 | 0 | 0 | 0 |
| soleirolii | 0 | 18 | 27 | 18 | 0 | 9 | 18 | 18 | 9 | 0 |
| tenorea | 0 | 17 | 83 | 100 | 0 | 17 | 17 | 17 | 17 | 0 |
| truncatula | 0 | 3 | 71 | 73 | 0 | 1 | 2 | 2 | 1 | 0 |
| turbinata | 0 | 6 | 74 | 79 | 0 | 3 | 9 | 7 | 3 | 0 |
CHR=chromosome number; ANT= anthracnose; GHAB=growth
habit; WVIG= winter vigor; COIL= Coils on pods; DAYSP=Days
to full pod production; HTH=Plant height; SPRD= Plant maximal
spread; SPIN= Pod spines; SCUR=Spine curvature
| Phenology | Production | |||||
|---|---|---|---|---|---|---|
| DAYSF | SDWT | BMASS | BMSP | GMPR | PODP | |
| arabica | 89 | 89 | 89 | 3 | 80 | 89 |
| blancheana | 83 | 100 | 83 | 50 | 72 | 83 |
| brachycarpa | 0 | 60 | 0 | 0 | 0 | 0 |
| ciliaris | 77 | 92 | 77 | 9 | 64 | 77 |
| constricta | 69 | 96 | 69 | 6 | 50 | 69 |
| coronata | 100 | 92 | 100 | 8 | 96 | 100 |
| disciformis | 100 | 100 | 100 | 8 | 96 | 100 |
| doliata | 89 | 91 | 89 | 3 | 70 | 88 |
| var. doliata | 33 | 0 | 33 | 0 | 33 | 33 |
| var. muricata | 35 | 95 | 35 | 0 | 23 | 35 |
| fischeriana | 40 | 80 | 40 | 0 | 20 | 40 |
| granadensis | 100 | 100 | 100 | 23 | 100 | 100 |
| heyniana | 50 | 100 | 50 | 50 | 0 | 50 |
| hybrid | 22 | 6 | 22 | 0 | 22 | 0 |
| intertexta | 94 | 88 | 94 | 29 | 71 | 94 |
| italica | 71 | 63 | 71 | 10 | 64 | 70 |
| laciniata | 85 | 98 | 85 | 9 | 81 | 85 |
| lanigera | 100 | 100 | 100 | 100 | 100 | 100 |
| lesinsii | 89 | 100 | 89 | 11 | 89 | 89 |
| littoralis | 45 | 95 | 45 | 3 | 40 | 45 |
| lupulina | 53 | 61 | 53 | 8 | 44 | 53 |
| medicaginoides | 0 | 100 | 0 | 0 | 0 | 0 |
| minima | 88 | 89 | 88 | 2 | 84 | 88 |
| monantha | 32 | 61 | 32 | 0 | 32 | 32 |
| monspeliaca | 17 | 58 | 17 | 0 | 17 | 17 |
| murex | 87 | 94 | 87 | 7 | 83 | 87 |
| muricoleptis | 88 | 100 | 88 | 0 | 63 | 88 |
| noeana | 100 | 89 | 100 | 11 | 95 | 100 |
| orbicularis | 65 | 75 | 66 | 3 | 55 | 65 |
| polyceratia | 0 | 100 | 0 | 0 | 0 | 0 |
| polymorpha | 79 | 47 | 79 | 2 | 39 | 79 |
| var. brevispina | 91 | 100 | 91 | 20 | 86 | 91 |
| var. polymorpha | 81 | 98 | 81 | 14 | 64 | 81 |
| var. vulgaris | 69 | 69 | 69 | 8 | 69 | 69 |
| popovii | 0 | 100 | 0 | 0 | 0 | 0 |
| praecox | 83 | 91 | 83 | 9 | 83 | 83 |
| radiata | 55 | 100 | 55 | 36 | 18 | 55 |
| rhodopea | 50 | 0 | 50 | 0 | 50 | 50 |
| rigidula | 83 | 85 | 83 | 3 | 78 | 83 |
| rigiduloides | 79 | 99 | 79 | 2 | 66 | 79 |
| rotata | 86 | 76 | 86 | 29 | 67 | 86 |
| rugosa | 88 | 88 | 88 | 26 | 79 | 88 |
| sauvagei | 100 | 100 | 100 | 40 | 100 | 100 |
| scutellata | 89 | 97 | 89 | 31 | 77 | 89 |
| secundiflora | 100 | 100 | 100 | 0 | 50 | 100 |
| shepardii | 100 | 100 | 100 | 0 | 100 | 100 |
| soleirolii | 18 | 100 | 27 | 18 | 18 | 27 |
| tenorea | 100 | 100 | 100 | 17 | 100 | 100 |
| truncatula | 74 | 55 | 74 | 3 | 69 | 74 |
| turbinata | 81 | 84 | 81 | 9 | 76 | 81 |
DAYSF=Days to flower; SDWT=100 seed weight; BMASS=Biomass among species; BMSP= Biomass within species; GMPR=Grams of seed produced ;PODP=Visual pod production
Objectives
• Develop a national GPP to serve as a source of new recombinants among the total diversity of germplasm available. This pool potentially would provide recombinants not obtained in the more genetically restricted regional pools described below.
• Develop regional GPP's that are adapted to each of four regions in the USA. Under the typical management practices for alfalfa in a particular region, the regional pools should be agronomically more acceptable than the national GPP. They also should have at least moderate levels of the traits available in varieties currently used in that region, plus genes that may be used to improve cultivated alfalfa.
Procedures
STEP 1: Bulk 100 seeds from each of the perennial PI's in the collection.
STEP 2: Plant 0.1 acre in field isolation to produce Syn. 1 seed. Three adjacent blocks should be planted and cut at different times to enhance overlap of pollination among the PI's in the regrowth.
STEP 3: To promote genetic recombination, produce Syn. 2 seed in field isolation. Fifty pounds of seed should be adequate to supply alfalfa researchers with a sample of the GPP. Seed should be produced in the Southwest to prevent natural selection for winter survival if a second year of production is necessary.
Four regions are suggested for developing GPP's: Northern, Intermountain Dryland, Southeast, and Southwest. Within each region, developing several pools organized by genetic and/or geographic origin would enhance opportunities for heterotic recombination in controlled cross-pollinations. Resource constraints, however, probably will allow for only a single GPP per region. The overall plan is to evaluate the PI's within the region where they are predicted to be most adapted according to the Minnesota research (Basigalup, 1989). Plants will be selected based on field performance, then crossed onto a bulk of current varieties used in that region to allow recombination of genes from the PI's with those from more adapted, elite backgrounds. Germplasm pools created in this manner should be simple to develop, while containing most of the genes in current varieties plus genes from the PI's that may enhance the rate of varietal improvements. Further improvements in the GPP's could be done by individual breeders according to their own strategies. Detailed steps of this plan are illustrated and outlined on the following pages.
STEP 1: Predict adaptation of each of the PI's to specific regions of the USA
by using data from the perennial core subset collection (Basigalup, et al.,
1992)
STEP2: Establish regional committees consisting of breeders, plant pathologists, entomologists, plant physiologists, and other interested alfalfa scientists to determine specific objectives and procedures for developing and improving GPP's in each region.
STEP 3: Screen and select the most adapted plants. In each region, establish spaced-plant nurseries (18" plant spacing) to evaluate the PI's that were predicted to be the most adapted to that region. In most regions, the PI's can be assigned among two or more breeders to reduce individual effort and to allow for selection in various environments. Locations and management conditions should be typical for alfalfa production in that region. Number of years of evaluation should be decided by the regional committee. In the North, for example, evaluation should span one or two winters to assess winter survival.
Select the most vigorous and agronomically acceptable plants. Discarded plants will be considered too unadapted for more immediate use in varietal development. Remnant seed of these PI's might be useful for long-term backcross or biotechnical programs to incorporate useful genes into adapted backgrounds.
Two alternatives regarding selection method should be considered by the ACGC. The first is to phenotypically select plants from within each PI so that the GPP is equally represented by all PI's within that region. The second alternative is to select both among and within the PI's, similar to the "ear-to-row-to-ear" method described for corn (Lonnquist, 1964). Although this second type of GPP may not be as genetically variable at first, it probably will be more adapted. But consider this question: Does enough genetic variability exist within the PI's to justify the first type of selection?
STEP 4: Hybridize selected PI's with improved varieties. In an isolated field, cross-pollinate the selected plants to a bulk population of the most current varieties used in that region. This varietal bulk should consist of a number of varieties of diverse genetic background, and have a high level of expression (if available) for the economically important traits, such as pest resistance, in that region.
Backcrossing to the varietal bulk might be considered, but it might dilute the frequency of genes from the PI's to a level difficult for finding genes needed in the future.
STEP 5: Identify hybrids. Among progenies having the PI's as maternal parents, select plants with a pest resistance derived from the varietal bulk to identify progenies likely to be true hybrids between the PI's and the varietal bulk. Selecting for a simply inherited resistance, such as anthracnose (Race 1), would be easiest; especially if plants can be evaluated as seedlings.
STEP 6 &7: Promote genetic recombination. Transplant hybrids to an isolated field and allow intermating to produce Syn. 1 seed. Then plant Syn. 1 seed to an isolated field (about 0.5 acre) to produce about 50 lbs. of Syn. 2 seed. This Syn. 2 seed would be available for distributing to researchers.
Result: GPP(s) semi-adapted to each region and having multiple pest resistance and other traits at levels where breeders can sample the pool and select plants with desirable traits for developing varieties. Adaptation of this type of GPP should be good enough to be useful to breeders for developing varieties. It would also serve as a reservoir of genes in a semi-adapted background for initial screening to identify genes that may be necessary for future breeding.
Anticipated time frame for developing useful regional GPP's is 5 to 7 years, depending on the traits desired for each region.
One additional suggestion is to develop a PI GPP for each region by intermating only the selected PI's; i. e., without crossing with the improved varieties. These GPP's may mask or dilute desirable traits less than would the PI x Variety GPP's. When searching for new traits, a logical sequence is to screen current varieties, regional PI x Variety GPP's, PI GPP's, the national GPP, then individual PI's.
Every 5 years, PI's collected since developing the regional GPP's should be evaluated and selected as was done when developing the original pools. Selected plants should be crossed with the most improved varieties used in that region, followed by identifying hybrids and subsequently producing Syn. 1 and 2 seed as described in Steps 3-7 for the original GPP's. Syn. 2 seed will be regarded as the Addition GPP's according to Hanson et al. (1972) terminology.
At this point more ideal procedures have been proposed by researchers to add new germplasm and improve the GPP. We anticipate, however, that both funds and researcher motivation needed for large-scale efforts may be available only for developing the original GPP's. Therefore, a simpler proposal is to make the Addition GPP available to researchers for incorporating into their own improved versions of the original GPP. For breeders who have not acquired seed of the original GPP by the time that the Addition GPP is available, seed of the Addition GPP should be mechanically mixed with the original GPP proportionate to those PI's already in the pool. Because adaptation of this Addition GPP should be nearly equal to that of the original GPP, mixing higher proportions into the pool as proposed by Barnes et al. (1977) is not necessary to offset the disadvantage in adaptation. And by crossing the new PI's to the most improved varieties, genes for newer traits in varieties will be added to the GPP, although at low to moderate frequencies.
Time frame for developing Addition GPP's probably will be the same as for the original pools, but presumably with less effort because of fewer PI's.
Barnes, D. K., E. T. Bingham, R. P. Murphy, O. J. Hunt, D. F. Beard, W. H. Skrdla, and L. R.Teuber. 1977. Alfalfa germplasm in the United States: genetic vulnerability, use, improvement, and maintenance. USDA-ARS Tech. Bull. 1571.
Barnes, D.K., B.P. Goplen, and J.E. Baylor. 1988. Highlights in the USA and Canada. In: Alfalfa and Alfalfa Improvement. A.A. Hanson, D.K. Barnes, and R.R. Hill, Jr. eds. American Society of Agronomy Monograph Series. No. 29. Madison, WI pages 1-22.
Bauchan, G.R. 1999. Use of annual medics in sustainable agriculture systems. In: Lucerne and Medics for the XXI Century. Proceedings of the XIII EUCARPIA Medicago spp. Group Meeting. Perugia, Italy, Sept. 13-16, 1999.
Basigalup, D.H., D.K. Barnes, R.E. Stucker. 1995. Development of a core collection for perennial Medicago plant introductions. Crop Science 35:1163-1168.
Diwan, N., Bauchan, G. R. and McIntosh, M. 1994. A core germplasm collection for the United States annual Medicago germplasm collection. Crop Science 34:279-285.
Hanson, C. H., T. H. Busbice, R. R. Hill, and others. 1972. Directed mass selection for developing multiple pest resistance and conserving germplasm in alfalfa. J. Env. Qual. 1: 106-111.
Lonnquist, J. H. 1964. Modification of the ear-to-row procedure for the improvement of maize populations. Crop Sci. 4:227-228.
Prosperi, J. M.., M. Angevain, I. Bonnin, E. Chaulet, G. Génier, E. Jenczewski, I. Olivieri and J. Ronfort. 1996. Genetic diversity, preservation and use of genetic resources of Mediterranean legumes: Alfalfa and medics. In: The Genus Medicago in the Mediterranean Region: Current situation and prospects in Reseach. Hammamet, Tunisie. October 1995. Options Méditerranéennes 18:71-89.
Small, E., M. Jomphe. 1989. A synopsis of the genus Medicago (Leguminosae). Canadian Journal of Botany 67:3260-3294. Compilation of Recommendations
Genetic Vulnerability (page 8)
There is a need to broaden the germplasm base used in alfalfa cultivars released by the alfalfa seed industry. The introduction of wild perennial germplasm and very non-dormant types from the Saudi Arabian Peninsula are examples of the beneficial attributes available.
Enhanced support for germplasm evaluation and enhancement projects would encourage
the use of a broader germplasm pool by plant breeders which would lead to more
diverse alfalfa cultivars
Core Subsets
Perennials (page 19)
1. The perennial Medicago core subset should receive high priority for
increase, maintenance and evaluation of new traits, e.g. disease and insect
resistance, quality traits, bypass proteins, water use efficiency, etc.
2. Specialty collections should be evaluated such as the highly non-dormant alfalfas (Steve Smith), the Chinese M. ruthenica collection (Austin Campbell and Larry Teuber), and the VIR collection in Russia. A core should be developed from this material by evaluating these collections in comparison to existing perennial Medicago core entries from the same geographic proximity group or species. Based on these evaluations the perennial Medicago core subset should be increased to represent the same frequency as the original core, i.e. about 15 - 20% of the specialty collection.
3. Additional germplasm collections are needed for secondary species ("wild" non-sativa species) from southeast China (Himalayan Mountains), southwest China, and southern Russia (Caucuses Mountains).
4. The core subset should be expanded as the number of accessions in the collection markedly increases. The new entries should be grown out and evaluated in comparison to existing perennial Medicago core entries from the same geographic proximity group or species.
Annuals (Page 19 - 20)
1. The annual Medicago core subset should receive high priority over
the remaining collection for increase, maintenance and evaluation of additional
traits especially freezing tolerance, disease and insect resistance.
The annual Medicago core subset should be evaluated for freezing tolerance,
either to extend the autumn growth in the Mid-West or for over-wintering in
milder climates such as California and south Texas.
3. The annual Medicago core subset should be evaluated for disease resistance such as Anthracnose, Phytophthora, Fusarium, Rhizoctonia, Pythium, Leptosphearulina, Phoma, and Rust. These diseases have been seen on some of the annual medics which have been grown in the U.S. If the annual medics are to be grown in the U. S., we need to know in advance if there are any disease problems.
4. The annual Medicago core subset should be evaluated for insect resistance to aphids and leafhoppers. These insects have been seen on some of the annual medics which have been grown in the U. S. If the annual medics are to be grown in the U. S., we need to know in advance if there are any insect problems.
5. The annual Medicago core subset should be evaluated for isozyme and molecular marker traits such as RFLPs, RAPDs, and microsatellites to further evaluate the collection for redundancies. The annual Medicago species are all self-pollinating and are thus relatively uniform within an accession.
6. No new collection trips are needed for the annual medics. There is an extremely large germplasm collection of 23,000 accessions located in Adelaide, South Australia. If additional accessions are needed of a particular species they can be easily obtained from this collection.
Rhizobium (Page 20 - 21)
1. A core subset needs to be developed using the data available in the GRIN system utilizing cluster analysis to identify subgroups. The final core subset should consist of between 65 to 75 isolates.
2. Once the core subset is established, the core should be evaluated for tolerance to low pH conditions. The isolates should be evaluated for both free living tolerance and symbiotic tolerance to low soil pH conditions.
3. Once the core subset is established, a determination of the symbiotic effectiveness of the core collection on the Medicago species core subsets should be conducted.
4. Any additional Rhizobium isolates should be sent to Dr. VanBerkum. Once a significant number of isolates has been added to the collection they should be evaluated in comparison to the core subset.
5. Evaluate the collection for Rhizobium strains by plant genotype interaction. There is a reasonable amount of information from other Rhizobia that there can be a substantial strain by cultivar origin. Since we recently identified strains of Rhizobia that will infect both alfalfa and soybean in China it seams likely that we could expect some significant interactions between genotypes from diverse locations. In a study that Don Barnes and I reported on we noticed some marked difference in the Rhizobia obtained from the same genotypes in different locations.
• Acquisition must be undertaken following guidelines arising from the Convention on Biological Diversity
• Gaps in the U.S. collection can frequently be filled by other collections around the world.
• The ACGC needs to establish guidelines that define when efforts warrant the acquisition of germplasm for inclusion in the U.S. collection, versus making use of germplasm that is available from other collections.
Primary Gene Pool
United States ( Page 23)
Given that funding can keep pace with a growing collection, the NPGS active collection should support the preservation of US cultivars that are no longer available through commercial sources. Although alfalfa is the fourth most widely grown crop in the US, NGO interest in conserving heirloom alfalfa varieties is poor to non-existent. Efforts need to be made to secure representatives of older varieties before seed stocks deteriorate. Materials registered with Crop Science should enter the active collection when the developer no longer distributes seed or it is not available from commercial sources. As non-CSR material is commercially retired, developers are encouraged to enter the obsolete cultivar into the NPGS collection. When PVP expires, cultivars should become part of the active collection, as PVP suggests the material is of exceptional value. At current levels of funding, a policy that preserves US cultivars as they are retired from commercial production cannot be implemented.
Cultivars of Other Nations (Page 23)
US users can make use of collections held in other institutes. An excellent source of information regarding the contents and availability of germplasm collections is the IPGRI Directory of Germplasm Collections http://www.cgiar.org/IPGRI/doc/dbintro.htm)
Areas where collection and preservation of cultivars may be beneficial to both the United States (based on limited representation in US collection) and corresponding countries (based on limited support for ex situ conservation, and lack of availability) include the Middle East, South Africa, and Mexico.
• Nondormant alfalfa continues to be underrepresented in the US collection. Efforts should be made to acquire the alfalfa germplasm collected by ICARDA during the 1970s, 1980s and 1990s.
Landraces and primitive cultivars (Page 24)
U.S users can make use of collections held in other institutes. Many of the
European nations contain extensive collections of local landraces and primitive
cultivars. For example, the European perennial Medicago database contains passport
data on 2888 accessions of 32 taxa, including 505 land race accessions stored
in 23 European genebanks http://www.cgiar.org/ecpgr/platform/ Crops/Medicago.htm)
.
Priority geographic areas recognized by the ACGC in 1988, and not yet collected
include the Himalayan region of India, south and northeast regions of China,
Sudan, Egypt and the Arabian Peninsula. Other geographic areas with limited
representation of landrace materials include South Africa, Northern Kazakhstan,
and adjacent Central Asian countries (i.e. Kyrgistan, Uzbekistan, Afghanistan),
Iran and Iraq.
Past CGC reports have placed emphasis on cultivated alfalfa that is adapted to acid soils and saline soils. There is increasing interest in developing alfalfa with increased water use efficiency, so drought tolerant germplasm would also be valuable to collect
Wild Medicago sativa ssp. sativa (Page 25)
Considering that the center of origin for Medicago sativa subsp. sativa
centers around Iran, Iraq, Azerbaijan and Turkmenistan, explorations to collect
wild material here, and in adjacent areas such as Southern Kazakhstan, Uzbekistan,
Afghanistan, and Tajikistan would be valuable, since wild-type M. sativa
subsp. sativa from this region are under-represented in the US collection.
• Efforts should be made to acquire germplasm collected during the 1980's
and 1990's through IPGRI-funded trips into Central Asia.
• Priority geographic areas recognized by the ACGC in 1988, and not yet collected include Georgia, Crimea and Southern Caucasus mountains
Medicago sativa complex (Page 26)
Fill the following geographic gaps in US collection through collection and donations. Species are listed in priority order:
1. Medicago sativa subsp. glomerata (2x=16), subsp. glutinosa
(2x=32) are grouped together since chromosome counts have not been obtained
to distinguish among the two subspecies: Southern Europe, Caucasus region, Algeria,
Tunisia
2. Medicago prostrata: Albania, eastern Austria, Greece, Czechoslovakia,
Hungary, Romania, Yugoslavia
3. M. sativa subsp. falcata: Northern Libya, Eastern Morocco,
Lebanon, Syria, Turkey, Central Asia (Turkmenistan, Afghanistan, Uzbekistan,
Tajikistan, northern Kazakhstan), Northeastern China, Northwest India, Northern
Pakistan, Southern Caucasus region, Ukraine, Northeastern Europe (Belarus, Lithuania,
Latvia, Estonia)
4. M. sativa nothossp varia: Northwest Iran, Syria, Northern,
Western, and Central Europe
Secondary Gene Pool (Page 26)
Fill the following gaps in US collection through collection and donations. Species are listed in priority order:
1. M. saxatilis: All areas of species distribution (see Appendix 1)
2. M. papillosa: Northern Armenia, Southern Georgia, Turkey
3. M. sativa nssp. tunetana: All areas of species distribution
(see Appendix 1)
4. M. sativa ssp. caerulea: Crimea, Central Asia
Tertiary Gene Pool (Page 27)
Fill the following gaps in US collection through collection and donations. Species are listed in priority order:
1. M. dzhawakhetica: Caucasus Mountains, Russia
2. M. daghestanica: Caucasus Mountains, Dagestan
3. M. rupestris: Lower Crimea Mountains, Ukraine
4. M. hybrida: Southern France
5. M. rhodopea: Southern Bulgaria
6. M. pironae: Alps of northeastern Italy
7. M. cancellata: Russia (North Caucasus, Caspian region)
Cultivated Medics (Page 27)
1. Most cultivated varieties of annual medic have been developed in Australia. Although they can be used in the southern latitudes of United States (notably California) they lack cold tolerance to more northern latitudes. Acquisition of accessions that are adapted to the northern boundaries of the species distribution may help us develop annual medic cultivars that can be used as cover crops or in ley farming in Washington, Montana and Wyoming.
2. The following cultivated species has been classified by Prosperi et al. as being vulnerable due to lack of representation in ex situ collections: M. arborea, M. italica, M. littoralis, M. rugosa, M. scutellata. Priority should focus on obtaining complete ecogeographic coverage of these taxa.
Non-cultivated Medicago species (Page 27)
Perennial species (Page27)
Efforts should be made to collaborate with other PGR institutes to conserve these resources in gene bank collections and in natural reserves.
Annual species (Page 28)
• Efforts should focus on determining if all accessions of the Lesin's collection should receive PI numbers. We need to collaborate with other PGR institutes to conserve those annual species that have potential use, or are vulnerable to genetic erosion.
• Efforts should be made to collaborate with other PGR institutes to conserve these resources in gene bank collections and in natural reserves.
Evaluations
Perennials (Page 28)
Efforts to verify ploidy have focused on M. sativa subsp. falcata, and M.
sativa subsp. caerulea. Work needs to be done to confirm ploidy status of the
Medicago sativa subsp. sativa collection
Approximately a third of the Medicago sativa subsp. sativa collection has been
evaluated for disease, insect, growth and morphological descriptors. To fully
utilize the collection, efforts need to focus on evaluating the remaining two
thirds of the alfalfa accessions. This will not be possible without increasing
the baseline budget of the NPGS Forage Legume CRIS.
Annuals (Page 28 - 29)
• Data on only one disease descriptor has been gathered, and for only the core collection, thus, additional diseases need to be screened.
• Some morphological descriptors have been taken, however, the majority of the accessions in the collection do not have even basic morphological descriptors. Thus a complete set of morphological descriptors needs to be obtained.
Over - all (Page 29)
Efforts should focus on acquiring a complete set of descriptors on all accessions,
especially all alfalfa accessions. A complete data set would allow users to
understand the diversity within the collection, and enable users to fully use
the collection.
The currently established core subsets should be re-evaluated and adjusted once
descriptor data has been obtained on the entire collection.
Accession classification based on molecular markers needs to be compared to
classifications based on morphological and ecogeographic descriptors. Can we
identify a combination of descriptors (that are easily and efficiently obtained)
that group accessions within the collection into meaningful clusters? Can we
establish a method to routinely classify newly acquired accessions to ensure
the collection continues to reflect a wide range of diversity but with a manageable
number of accessions?
Evaluations directed at traits that can broaden the use of alfalfa need to be
carried out. For example, there is current interest in the use of alfalfa as
a medicinal herb, for phytoremediation, use as biofuels, cover crop, etc. The
production of organic hay is increasing. The genetic variation within the germplasm
collection for these uses has been minimally explored .
Accession evaluation and observation data needs to be acquired and placed into the GRIN database in a format that allows users to draw valid comparisons across all accessions evaluated for that trait.
Enhancement (Page 30 and Appendix 4 Page 45)
1. Develop a national GPP to serve as a source of new recombinants among the total diversity of germplasm available. This pool potentially would provide recombinants not obtained in the more genetically restricted regional pools described below.
2. Develop regional GPP's that are adapted to each of four regions in the USA. Under the typical management practices for alfalfa in a particular region, the regional pools should be agronomically more acceptable than the national GPP. They also should have at least moderate levels of the traits available in varieties currently used in that region, plus genes that may be used to improve cultivated alfalfa.
3. An ARS alfalfa research geneticist position was established in 1999 at Prosser, WA. with an emphasis to carry out additional germplasm enhancement work. Enhancement work by this position should continue to be encouraged and supported.
Collection Preservation (Page 31)
Original seed needs to be inventoried with seed counts. Germination data on original seed lots needs to be obtained on all accessions with sufficient seed. Original and regeneration seed lots need to be stored at –15 to –20 C.
Germination data needs to be obtained on increased material to establish a
baseline to monitor seed viability and to provide a check to verify that regeneration
procedures are high quality.
• Research needs to be carried out to verify if our standard plant population
size during regeneration is adequate for limiting genetic drift, and if genetic
change in accessions is minimized during the standard regeneration process.